 魚が陸上を歩くまで
魚が陸上を歩くまで
第3章―5 感覚系
生存に必須な感覚系の陸棲化の過程は、骨の変化を追える中耳の耳小骨の形成以外はあまりはっきりしない。
第3章-5-1 視覚
脊椎動物の最も初期とされるHaikouichthysに既に双眼の存在が確認されるが(Shu et al. 2003)(Shu et al. 1999)、一般に軟組織である目は多くの場合化石として残りにくいため報告は少ない(Young 2008)。従って、視覚・目の進化の過程を知ることは困難である。そのため系統樹のさまざまの位置にある現生種の目の詳細研究から推し量ることが試みられている。
目形成の主要遺伝子とされるPax 6の起源は海綿動物が分岐する前に起こった遺伝子重複まで遡る。その遺伝子重複後有棘胞類において網膜で発現するPax Aや目柄:eye stalkで発現するPax BやPax Eが生じる。そしてその後に出現した左右相称動物において、目の形成に関係するPax 6が生じたとされる(Suga et al. 2010)。従来は系統ごとに目が数十回も独立に平行進化したと考えられた。しかし、目形成のPax 6やSix 1, Six 3遺伝子がプラナリアからヒトまで共通であることから、現在では目の起源は単系統であったと考えられている(Gehring 2005)。Pax 6、Eya1、Six1をマスター制御遺伝子として種々の形態形成遺伝子が入り込んで、系統ごとに異なる目形成のネットワークが作り上げられてきたと考えられる(Collin 2009)。
明暗を識別する器官、即ちnon-image forming eyesと物を見ることが出来る目、即ちimage forming eyes:カメラ目では機能、構造はもとより受光細胞にも大きな違いがある。脊椎動物の系統では、ヌタウナギはホヤのようなnon-image forming eyesであるが、ヤツメウナギは、その幼体がnon-image forming like eyesであり、変態した成体がimage forming like eyes を持つ。ヤツメウナギより系統樹で上位にある軟骨魚類と硬骨魚類のいずれの目もimage-forming 型なので、ヤツメウナギはnon-image forming eyesとimage-forming eye の橋渡し的な位置にある(Collin et al. 2009)。
受光細胞は5系統あり、明順応:photpic:bright light visionのcone-based cellと暗順応:scotopic:dim light visionのrod-based cellに大別され、前者はさらに感応する色によって分けられる。従って、色覚は網膜上に配置された受光細胞を詳細に調べることで推定できる。
ヤツメウナギの分子解析から、5系統の受光細胞は顎口類と無顎類が分岐するまえに起源を持つことが明らかにされている。即ち、700~540Myaの間にcolor vision が既に獲得されていたことになる(Collin 2009)。光受容体オプシンの進化系統樹を図1に示す。

無顎類と顎口類の共通祖先は4つのオプシン:LWS, SWS1, SWS2, Rhを持つと仮定(Collin et al. 2003)。Rh:無顎類はRhAとRhBを、顎口類はRh1とRh2を持っているので、各々独立に遺伝子重複があったと考えられる。主要な5種:LWS, SWS1, SWS2, Rh1, Rh2は、条鰭類、ハイギョ、両生類と一部の羊膜類で保存されている。但し、条鰭類ではさらに遺伝子重複3Rを受けてLWS2, SWS2A, SWS2B, Rh2Aα, Rh2Aβが加わっている(Perry et al. 2005)。板鰓亜網ではRh1以外のオプシンの存在は不明。シーラカンスではLWS, SWS1, SWS2 が失われている。 From Collin 2009
オーストラリアの現生ハイギョNeoceraodus forsteriは、デボン紀における肉鰭類と似た目を持っていると考えられる(Collin et al. 2009)(Bailes et al. 2007)。ハイギョの網膜にある受光細胞と色素を詳細に解析した結果、幼体では紫外線域までの色覚を持つが、成体ハイギョの色彩感覚は魚類や両生類よりもカメや鳥類に近いと判断された(Hart et al. 2008)。また、幼体が紫外線域までの色覚を持つのは食性が関係しているとされる。成体はカタツムリや貝を食べるが、幼体は動物性プランクトンを餌としており、紫外線を吸収、ないし反射する動物プランクトンに対する視覚確保のためであろうとされる。成体はより深いところでも生活するようになるので紫外線依存が減るのであろう(Keler et al. 200)。
陸棲化に関与した肉鰭類の受光細胞もハイギョのそれと大差ないものであったろうと思われる。
最近板皮類の眼球の運動を担う筋肉に関する情報が化石から得られている(Young 2008)。それによると初期顎口類である板皮類は、現生ヤツメウナギと同様7本の互いに拮抗する機能を持つ筋肉系で眼球運動を行っていたことが明らかになった。即ち、初期顎口類は既にimage forming eyes を獲得していたし、眼球運動を担う筋肉系や神経系も形成されていた。なお、現生脊椎動物では6本の筋肉系で眼球運動を行っている。
これらのことから、肉鰭類が陸棲化にするにあたり、色彩感覚や目の構造を含め目の基本的な機能は既に備わっていたが、水中で機能する目から大気中で機能する目になるためには、目に入る光量の増加、異なる屈折率を持つ媒質、目の乾燥への対応が必要であった。
光量増加への対応
水中より大気中の方が太陽光強度が強いので、陸棲化に際しては網膜を保護するために目に入る光の強度を下げる必要があった。
脊椎動物は魚類の段階から受光細胞内にフィルター機能を持つoil droplets を持っていたとされる(Robinson 1994)。ハイギョの網膜には、図2に示すように1種類のrod状受光細胞と4種類のcone状受光細胞が存在する。4種のcone状受光細胞のうち色素を持たない3種はoil dropletsを含有している。こういったoil dropletsの違いは昼行性や夜行性といった生態と関係している。現生のトカゲ類、鳥類の受光細胞も似た構成になっているが、現生の両生類や哺乳類、一部の夜行性爬虫類ではoil dropletsが減少している。
なお、受光細胞中のoil dropletsは、目の分解能と感度という互いにトレード・オフの関係にある特性に関連しており、生息場所や採餌行動に適化させるように進化したと考えられる (Hart et al. 2008)。

R:rod:桿状体、C:cone:錐状体、dm:distended mitochondoria:巨大ミトコンドリア、m:mitochondria within the inner segment:内節にあるミトコンドリア、 n:nucleus:核、od:oil droplet:油滴、os:outer segment:外節、p:paraboloid、yp:yellow pigment:黄色素 From Collin et al. 2009
ハイギョの色覚に関するこれらの結果から、初期四肢類の色覚は浅い水辺で獲得されたものであり、肉鰭類がデボン紀に上陸したときには既によく発達したcolor visionを進化させていたと類推される(Hart et al. 2008)。太陽光は水中では急激に減衰するので、浅瀬に進出した385Mya頃に光強度の増加への対応が始まったと思われる。フィルター機能を持つoil dropletを増やすことによって容易に、従ってかなり短期間で対応出来たと考えられる。
異なる屈折率を持つ媒質への対応
大気と水では光に対する屈折率が大きく異なるので、陸棲化に際しては水晶体を変える必要があった。水晶体の重要な構成要素としてクリスタリンがある。魚類のクリスタリンは硬度があり、屈折率が大きく、水分含有量が少ないが、陸棲化に伴い柔軟性のある、屈折率の小さいクリスタリンに変化している(Wistow et al. 1993)。
α、βクリスタリンはほぼ総ての脊椎動物に共通して存在しているが、γクリスタリンは多様化しており系統ごとに異なっている。従って、γクリスタリンの遺伝子は各種脊椎動物の系統樹における位置づけに有用である。γMクリスタリンは魚類に特有であり、γA~γFクリスタリンは魚類にはホモログが見られないので陸棲化に伴って生じたと考えられる(Wistow et al. 2005)。
こういった特に適応的なγクリスタリンを選択することにより水晶体の屈折率を下げることは出来たが、クリスタリンの屈折率を変えるだけでは大気と水の間の大きな屈折率差を解消するには不十分である。そのため主にレンズの厚さを変えることで陸棲化に対応してきた。即ち、魚類の分厚い所謂魚眼レンズを陸棲化に際して薄いレンズに変えている。
空気に比べて屈折率の高い水中にすむ硬骨魚の目のレンズは、眼窩という限られた空間の中で焦点を結ぶために分厚い魚眼レンズになっている。網膜上に焦点をあわせるには、水晶体と水との屈折率差が小さいためレンズの厚さを変える方式ではなく、レンズを水晶体牽引筋と呼ばれる筋肉で前後することで調整している。
陸棲化に伴い毛様体筋によってレンズの厚さを変える焦点調節方式に進化したと考えられるが、水晶体牽引筋から毛様体筋に、いつどのような過程を経て進化したのかは不明である。Tiktaalikはその前肢で状態を支えることが出来たとされ、大気中で機能する目を持っていた可能性があり、屈折率への対応は383Mya頃に始まったと思われる。昆虫などを捕食していたと考えられる小型の四肢類が出現した340Mya頃には、目によって昆虫を捕捉する必要があり、従ってある程度の焦点調節方式を進化させていたであろう。
乾燥への対応
脊椎動物の陸棲化に伴い目を乾燥から守るために瞼が進化した。両生類は上下に瞼を持ち、さらに目をおおう薄い透明な瞬膜(しゅんまく)がある。これらによって目の乾燥を抑えている。また、水分を補給するシステムも進化させている。眼窩腺がそれであり、両生類や爬虫類、鳥類では油性の液体が分泌される。哺乳類では眼窩腺の1種である涙腺が発達しており、水分が分泌される。これらのシステムがいつ、どの順番で、どのように進化してきたのかは不明である。両生類が瞬膜や瞼、眼窩腺を持っていることから、浅瀬に進出し曝気の機会が増えた365Mya頃に瞬膜や瞼、眼窩腺に向けた進化が始まったと思われる。
角膜上皮の微細構造が種々の環境に棲む様々の動物についてSEMで調べられている。角膜上皮細胞の数は平方ミリ当たり28860~2126個と棲む環境に依存して大きく変わることが明らかにされた。角膜上皮細胞の面密度は海生、淡水生、陸生の順に少なくなる傾向を示す。肉鰭類が陸棲化する時に角膜上皮細胞の面密度が急激に減少したとされる(Collin & Collin 2006)。海生種と淡水生種の違いは角膜表面にかかる浸透圧と関係すると思われる。耐乾燥性と関係している可能性があるが、角膜上皮細胞と耐乾燥性の関係は不明である。角膜上皮細胞の面密度を下げることはそれほど困難な適応ではなかったであろう。
目の物理的保護
目を水圧から保護するためにサメ類や一部の魚類は眼球の強膜に軟骨や骨の組織、ocular skeleton :OSを持っている。軟組織の目は化石化しにくいが、このOSは化石化しやすく、目の進化の一部を探ることが可能である。
顎口類の初期とされる板皮類の1系統の節頸類ではOSは4個からなり、初期条鰭類も4個の骨からなる。肉鰭類では20個以上の小骨からなる。サメでは初期は肉鰭類よりさらに多数の小骨からなるリング状であったが、現生種では1個のカプセル状へと大きく変化している。古生代のサメのOSは現生爬虫類のそれに似ており、現生爬虫類のOSは焦点調節機能を持つので古生代のサメや肉鰭類も焦点調節機能を持っていた可能性がある(Pilgrim et al. 2009)。しかし、魚類のOSと現生爬虫類のOSは相同器官ではないとされ(Franz-Odendaal et al. 2006)、古生代の軟骨魚類の焦点調節機能に関してははっきりしない。なお、OSは強膜骨板:Sclerotic ring とも呼ぶ。

軟骨魚類の例として化石種と現生種を、Sarcopterygiiの例として現生爬虫類を示してある。化石軟骨魚類と現生爬虫類では似たようなOSであるが、相同でない可能性が大きい。
From Pilgrim et al. 2009
第3章-5-2 聴覚
耳は姿勢や加速度を検知する平衡感覚機能と音を感知する聴覚機能を持っているが、前者の機能は脊椎動物の初期段階に既に獲得しており、後者の機能も陸棲脊椎動物とは異なるもののやはり脊椎動物のかなり初期段階で進化させていた。
哺乳類の耳は外耳、中耳、内耳の3部分からなる。音を集める外耳は哺乳類になってから獲得したものであり、必要な音波を効率よく捉えるように進化してきた。外耳で捉えた音波は外耳道を通り中耳に導かれる。中耳は陸棲化に伴って大気中における聴覚を可能にするために獲得された部分であり、大気中を伝わる音波振動を鼓膜の振動に拡大変換する機能を持った耳小骨からなる。内耳は魚類でも哺乳類でも大別すると平衡感覚部と音の検出機能を持つ聴覚部とからなる。クレード間、あるいは種間での聴覚部分の違いは大きいが、平衡感覚部は似たような構造である。しかし、内耳の2つの機能部分を構成する器官は同じ原理に基づいた似たような基本構造になっている(Popper & Fay 1997)(Fay & Popper 2000)。即ち、リンパ液で満たされた嚢があり、その中には有毛の神経細胞が多数配列されていて、これら神経細胞が感覚毛の動きで情報を検出出来るようになっている。
内耳の平衡感覚部は、陸棲脊椎動物や多くの現生魚類では3つの管(三半規管)からなり、空間を3次元で感知できるようになっている。この部位は行動の活発化とともに進化してきており、ヌタウナギでは1つの管だけで形成されていたが、原始的な魚類の段階で2つの管を持つようになり、顎口類の段階で3つの半円からなる三半規管を持つ内耳を獲得した。内耳の平衡感覚を受け持つ三半規管部分は陸棲化に際してあまり大きな変化を要しなかった。
一方、内耳の音波を捉えるラゲナと呼ばれる聴覚部分は、陸棲化に際して内耳が収まっている頭蓋骨を含めて大きな変化をしている。ラゲナは円口類にも既に見られるが、魚類はもとより両生類の段階になっても内耳の中でそれほど大きな部分を占めてはいないし、カメといった爬虫類の一部でもそれほど大きくはない。しかし、ワニや鳥類、哺乳類では長大化しており、特に哺乳類では蝸牛管と呼ばれる渦巻状の長い管になっている。魚類は体表にあたった音波を骨伝導により直接内耳のリンパ液で満たされたラゲナに伝え、生じたリンパ液の振動を感覚毛の変形により感知する。一方、陸棲脊椎動物では、空気の振動を鼓膜の振動として受信し、鼓膜の振動を中耳を介して内耳の聴覚部分に伝える方式に変えている。そのため陸棲化に際して内耳の聴覚部は大きな変化を要したのであろう。
脊椎動物は陸棲化に際して、元来は呼吸用であった第1鰓孔の外表面に鼓膜を形成し、鰓孔を中耳に変えている。中耳には耳小骨と呼ばれる小さな骨があり、この骨で鼓膜の振動を内耳の聴覚部に伝えるシステムを進化させた。
両生類も羊膜類も耳小骨によって鼓膜の振動を内耳に伝えている。哺乳類の系統では鐙骨と槌骨、砧骨の3個の小さな骨で中耳を構成しており、梃子の原理で音を拡大する機能を持つ。両生類と爬虫類の中耳は耳小柱ただ一個からなる。これらの骨の起源に関しては完全に解明されたわけではないが、化石記録と胚発生観察とから一応の結論が出されている(シュービン)(Clack 2001)。鐙骨は第二鰓弓を起源としており、軟骨魚類を含めた魚類の舌顎骨に、そして爬虫類が唯一つ持つ中耳骨の耳小柱(鐙骨)に対応している。槌骨と砧骨は第一鰓弓を起源としており、各々魚類の関節骨と方形骨に対応しており、爬虫類では下顎を構成する骨になっている。中耳は魚類が上陸したときにただ1回進化したと考えられたこともあったが、耳小骨の構成の大きな違いから、哺乳類と両生類や爬虫類は独立に鼓膜つきの耳を進化させてきたと考えられている(Manley 2010)(Koppl 2009)(Meng et al. 2011)。
舌顎骨は条鰭類や肉鰭類にとって顎の開閉や鰓蓋の開閉に必要なものであり、摂食活動や呼吸活動に関与する重要な機能を持っていた。魚類から肉鰭類を経て四肢類へと移行するなかで、水流とともに餌を飲み込む方式から噛み付く方式へと摂食方式が変化し、舌顎骨としての機能が低減した。さらに曝気の機会が増えるに伴い鰓機能の衰退と肺呼吸への移行が起こり、舌顎骨としての機能を失った。四肢様魚類のパンデリクスの段階で舌顎骨は急激な再配置を行い、イクチオステガでは依然として水中で機能する耳であったが、既に鐙骨は聴音機能を持っていたと考えられている(Clack 2006)(Clack et al. 2003)。イクチオステガよりは陸棲化していたとされるアカントステガの段階で顎を構成する骨から頭骨の一部に組み込まれた(ジンマー) (Brazeau & Ahlberg 2006)。しかし、アカントステガでも側線系を保持しており、初期両生四肢類段階の耳の聴覚機能は現生肉鰭類や現生の原始的な条鰭類のそれに近かったであろうと考えられる(クラック)(Clack 2001)。初期両生類のアカントステガ、グリールペトン、さらには初期羊膜類とされるエンボロメリでも舌顎骨は、やや扁平で魚類のそれより小さくなっていたが、まだオステオレピスの舌顎骨に似た状態で口蓋の骨に関節している。また、耳周辺の構造、仕組みいずれもオステオレピスに似ており、たとえ大気中で聴覚が機能したにしても狭い音域しか捉えることが出来なかったと予想されている。
肉鰭類から四肢類へと移行するとともに頭部を構成する骨の間の結合が強化され、その結果舌顎骨が頭骨に組み込まれた状態から外れ、聴覚の方に振り分けることが出来たのは、石炭紀になってからである(ジンマー)。なお現生両生類では幼生時期には側線系を保持しているし、終生水棲である両生類だと成体でも側線系を持つが、羊膜類は胚の段階でも側線系の痕跡すらみられず、耳が完全に聴覚機能を受け持っている。
四肢様魚類から四肢類に至る耳小骨の変化を図4に示す。
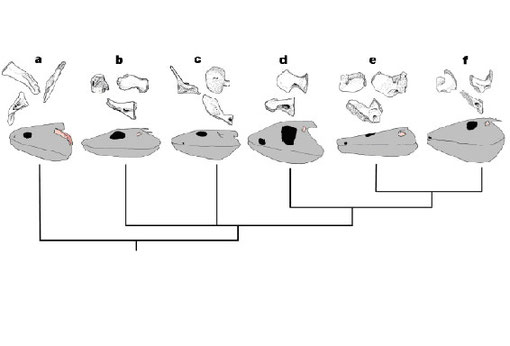
a:Eusthenopteron(デボン紀後期), b:Acanthostega(デボン紀末期), c:Ichthyostega(デボン紀末期), d:Pederpes(石炭紀初期), e:Greererpeton(石炭紀中期), f:Pholiderpeton(石炭紀後期) 灰色:頭蓋骨 ピンク:舌顎骨/鐙骨 上左:側面 上右:背面 下:後面
From Clack et al. 2003
魚類の舌顎骨は初期四肢類の段階で縮小し、大気中で機能する中耳の耳小骨へとその機能を変えたが、この骨の聴覚への適応はそれほど急激ではなかったようである。まともな鼓膜つきの耳が最初に現れるのは、石炭紀初期ヴィゼアン世のバラネルペトン :Balanerpetonであり、鐙骨の配置などは現生カエルのそれに似ている。
舌顎骨が縮小したことにより、その周辺の骨と筋肉の役割にも変化が生じ、その結果頸が形成され、大きな肩関節が形成される空間が確保された。このような一連の連動した変化により四肢化が強化されたといえる。
さらに、頭蓋骨にも変化が必要であった。陸棲脊椎動物の内耳は頭蓋骨内に収まっているので、頭蓋骨に前庭窓と呼ばれる孔を形成し、中耳から内耳に振動を伝えるようにする必要があった。前庭窓から入った振動はラゲナ、又は蝸牛管内のリンパ液に伝わり、リンパ液の振動による感覚毛の変形により音が感知される。さらに前庭窓から入った進行波が戻り波となり、ノイズを発生するのを防ぐために蝸牛窓と呼ばれる小さな孔を頭蓋骨に形成した。
こういった大きな変革が必要であったために、バラネルペトンのように陸上で機能するそれなりの耳を獲得するまでに、肉鰭類が潟などの浅い潮汐域に進出してから55Myほど、初期四肢類が出現してからでも40Myほどの時間を要している。
無顎類の多くや魚類では頭部から尾部にかけて側線と呼ばれる感覚器官がある。側線は肉鰭類も保持しており、初期四肢類のアカントステガやイクチオステガにも見られる。また、現生両生類の幼生や水棲の現生両生類にも存在している。即ち、側線は周囲の水流や水圧を検出する水棲における必須器官である。
側線に沿って感丘(かんきゅう)や管器と呼ばれるリンパ液で満たされた嚢があり、内部に感覚毛を持った神経細胞が配列していて、基本的には内耳に見られる感覚器官の構造と似ている。有毛細胞は脊椎動物の初期形態とされるナメクジウオにも見られ、化石記録からも非常に古い感覚器官であることが分かる。また、内耳でも感丘でもPax2と呼ばれる遺伝子の発現が見られる。
こういったことから側線と耳は同じ原理に基づいた似たような構造であり、系統発生的には側線器官の一部が内耳に変化したものと考えられる。しかし、側線系と内耳は独立に進化したとする説もあり(Herbrand et al. 1998)、結論は出ていないようである。
耳と関連するプラコードは脊椎動物の初期段階で出現したとされる(Streit 2001)。Pax2は蝸牛管の成長に関与する遺伝子とされ、魚類でも哺乳類でもこのプラコード部でPax2やPax8の発現が見られる(Streit 2001)(Fritzsch et al. 2007)。また、中胚葉からのEGF-19シグナルや外胚葉の神経板:neural plateからのWnt8cシグナルも耳誘発に関与するとされており(Ladher et al. 2000)、平衡機能を持つ器官の起源は、耳と関連するプラコードが形成される前の左右相称動物が出現したころまで遡ることが、遺伝子解析からも確認されている(Herbrand et al. 1998)。
第3章-5-3 嗅覚と味覚
魚類では外鼻孔を1対しか持たないのもあるが、多くの魚類や軟骨魚類では鼻孔が隔壁によって前後に分割された2対(4孔)になっていて、前方の2孔が入水孔、後方の2孔が出水孔になっている。この構造により遊泳中は連続的に水が鼻腔内を流通し、匂い分子を捉えることが出来るようになっている。
魚類の鼻孔は臭覚器官としての機能しか持たないが、陸棲脊椎動物では鼻孔は口腔内に開いており、臭覚器官と呼吸器官の機能を併せ持つ。口腔内に開く内鼻孔は395Mya頃棲息していた肉鰭類Kenichthysにおいて獲得されている(Janvier 2004)。Kenichthysの内鼻孔は口腔先端部にあるが、365Mya頃棲息していたアカントステガでも似たような位置に留まっている。従って、アカントステガ段階でも空気は口から飲み込んでおり、鼻孔を呼吸には使っていなかったとされる(クラック)。扁平な頭骸骨と口吻はそのような呼吸方式に適応した形状とされる。アカントステガの鼻孔が、臭覚器官として大気中で機能したのかどうかは不明とされるが、水棲主体の生活であったことから大気中ではまだ臭覚期間としても機能していなかった可能性が高い。陸棲化に際して鼻孔が臭覚器官として大気中で機能し、さらに呼吸機能を持つようになったのはアカントステガよりはかなり後とされる。恐らく「ローマーの空隙」後の頭骸骨や口吻が扁平でない四肢類、即ち羊膜類とされる幅の狭い頭部を持ったウェエストロティアナが出現した頃に鼻孔は呼吸にも使われ始めたと考えられる。そして大気中で臭覚器官とし機能したのは、呼吸機能よりも先であったであろう。
内鼻孔は陸棲脊椎動物の特有の器官とされるが、完全な水棲であったKenichtysの段階で内鼻孔化が始まった理由は明らかにされていない。恐らくKenichtysが出現した頃に肉鰭類の多くが遊泳型から底生型に変ったと考えられる。底生化に伴い匂い分子を含んだ水が鼻孔内を流れなくなり、強制的に鼻孔内に水を流す必要から内鼻孔が形成されたと考えられる。即ち、鰓蓋を働かせることにより、外鼻孔から水を取り入れ、内鼻孔を通じて口腔内を経由して鰓から排出することで、停止状態でも嗅覚を機能させることが出来るようになったのであろう。
鼻腔の嗅粘膜の嗅上皮には嗅覚レセプターを持つ嗅細胞が並んでいる。魚類は線毛性嗅細胞、微絨毛(ビジュウモウ、ビニュウモウ)性嗅細胞、陰窩細胞の3種の嗅細胞を持つ。陸棲脊椎動物の嗅上皮には線毛性嗅細胞しかないが、鋤鼻器官を持つ場合にはは鋤鼻器官にある微絨毛性嗅細胞でフェロモンを感知している。嗅上皮は粘液に覆われていて、粘液の中に各嗅細胞は嗅毛を伸ばしている。匂い物質が粘液に溶け、嗅毛にある嗅覚レセプターに捉えられ、匂いとして感知される(岩堀 2011)。
臭覚レセプター:Olfactory receptorsには、鼻腔のOR:odorant, TAAR:trace-associated olfactory receptors、及び鋤鼻器官のV1R, V2R :vomeronasalといったファミリーがあり、いずれもG蛋白に結合したレセプターである。これらに関係した遺伝子の獲得は無顎類の時代まで遡るといわれる(Korcching 2009)(Niimura 2009b)。
臭覚レセプターORを哺乳類は1000個程度、硬骨魚類は100個程度持っており、魚類の方がORは少ないが、種間での違いは哺乳類より大きい(Niimura & Nei 2005)。ナメクジウオは嗅覚器官を持たないにもかかわらずORを30個以上は持っているが(Niimura 2009a)、ホヤなどの被嚢類ではORは見つかっていない(Raile et al. 2006)。
脊椎動物のORはtype 1とtype 2に大別され、さらにtype 1はclass Ⅰとclass Ⅱの二つに分けられる(Freitag, et al. 1998)。また、ORと形態的に似ているとされる遺伝子群:θ、κ、λは動物の系統ごとの違いはわずかであり、ORとは異なる遺伝子系であり、さらにκとλは嗅覚と無関係であるとされる(Niimura 2009a)。
脊椎動物のORをまとめる。
type 1;α、β、γ、δ、ε、ζ(class Ⅰ;α、β;class Ⅱ;γ)
type 2;η、θ1、θ2
硬骨魚類と四肢類の両方に最も近い共通祖先は少なくとも9種のORを持っていたが、その中の2つ、αとγは大気中で機能するORであり、四肢類系で多様化している(Niimura & Nei 2005)。ORの主要なグループであるβは水棲、陸棲どちらにも見られ、δ、ε、ζ、ηは水棲脊椎動物において機能している。現生シーラカンスや水棲の哺乳類では大気中で機能するclass ⅡのORを持つが、大部分が擬遺伝子となっている(Freitag et al. 1998)(Eisthen 1997)。ORに擬遺伝子が多いのは、環境との相互作用により容易に変化しうることを示しているが、同時にシーラカンスに見るように機能しなくても長期にわたり保存されており、進化の過程を追うのに向いている(Niimura 2009b)。
図5に種々のOR遺伝子の獲得・消失時期を示す。

四肢類の系統では、α遺伝子とγ遺伝子のグループが急激に数を増やしているが、陸棲化への対応のためと考えられる。 From Niimura 2009a
ORは特に陸棲脊椎動物で多様化しているが、同じく嗅覚上皮に見られるTAARは、魚類では13~109遺伝子を持つが、四肢類では3~22遺伝子しか持っていない。従って、TAARは魚類の臭覚で重要な機能を持っていると予想されるが、詳細はまだ明らかではない(Hashiguchi & Nishida 2007)(Hashiguchi et al. 2008)。図6にTAARファミリーの分岐状況を示す。TAARは5つのサブグループからなるが、この5つのグループは魚類と四肢類の分岐以前にすでに存在していた。しかし、その後の進化の過程で、四肢類への分岐後にサブグループⅢを失い、両生類はサブグループⅠを、羊膜類はサブグループⅤを失っている。また魚類においては全遺伝子重複WGDのためにTAARの進化の過程は複雑化している(Hashiguchi & Nishida 2007)。ORほど明確でないが、サブグループⅡとサブグループⅣが大気中で、サブグループⅢが水中で機能すると思われ、またサブグループⅠとサブグループⅤは水棲、陸棲どちらの脊椎動物にも見られる。
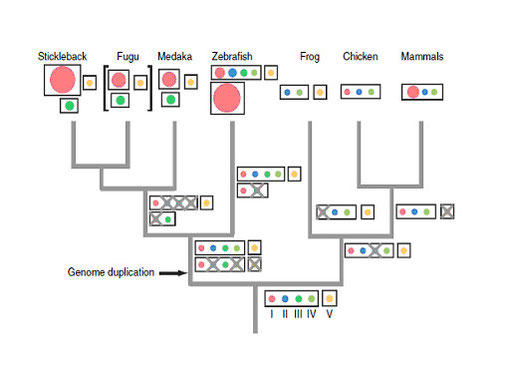
色付き円を囲む四角部:染色体に対応。色付き円:TAARの5つのサブファミリーで、円のサイズは各サブファミリーに属する遺伝子の数に対応。条鰭類と肉鰭類の共通祖先は5つのサブファミリーⅠ~Ⅴを持っていた。Ⅰ~Ⅳは同じ染色体上に、Ⅴは別の染色体上に存在。四肢類の共通祖先ではサブファミリーⅢが消失しているが、四肢類の系統では祖先のTAAR遺伝子の全体的な構造は保たれている。魚類の系統では、全遺伝子重複があったためにTAARファミリーは倍になっている。現生魚類のTAARファミリーの構造は系統ごとに異なり、複雑化している。 From Hashiguchi & Nishida 2007
かって鋤鼻器官は魚類には無く、脊椎動物が陸生化するときに獲得したとされた。しかし、両生類にも鋤鼻器官が見つかり、両生類と羊膜類の共通祖先が既に保持していたと考えられた。両生類と羊膜類の共通祖先は水棲なので、鋤鼻器官は上陸に伴って獲得されたのではなく、水棲時代に獲得されたものとなる(Jermakowicz et al. 2004)(Eisthen 1997, 2000)(Eisthen et al. 1994)。従って、鋤鼻器官はもともとは非揮発性の匂い物質の検出に用いられた器官であたったが、上陸時点で嗅覚細胞が二つに分かれたということらしい。
鋤鼻器官の臭覚レセプターV1RsとV2Rsは陸棲動物におけるフェロモンや普通の匂い物質の検出に関係している。V1RsとV2Rsも魚類と四肢類の共通祖先から引き継いだものであり、機能しているV1RsとV2Rsとの比は陸棲脊椎動物のほうが魚類よりも50倍ほど高い(Shi & Zhang 2007)。先のORのclass Ⅰとclass Ⅱの間でも水棲と陸棲でその数に大きな違いが見らており、V1RsとORのclass Ⅱ は揮発性の匂い物質に、V2RsとORのclass Ⅰは水溶性の匂い物質と関係しており、陸棲化に伴い鋤鼻機能が大きく変わったことを示す。
また、mouse とrat のV2Rsの特性にも大きな違いが見られ、鋤鼻器官の臭覚レセプターも棲息場所や生態等に適応してその数が系統毎や種毎に大きく変化していることが明らかにされている。鋤鼻器官の臭覚レセプターの特に上陸後の進化は顕著であり、ORレセプターとは異なった選択圧、恐らく性選択圧を受けて種ごとに独立に進化したと考えられる(Wenry & Zhang 2008)。
味覚器は接触刺激を、嗅覚器は遠隔刺激を検出する違いはあるが、いずれも化学物質を検出する受容器を持つ。脊椎動物は味蕾と呼ばれ口中にある味覚器によって味を感じている。現生円口類ではヤツメウナギは味蕾を持つが(Barreiro-Iglesias et al. 2010)、ヌタウナギは持っていないとされる(Braun 1998)(Finger 1997)。従って、味蕾は円口類の段階で獲得されたことが予想される。味蕾の構造と機能は他の感覚器官に比べると単純であり、原始的なヤツメウナギではやや異なるが、魚類、両生類から哺乳類まであまり大差ない。味蕾は口中の水分の多い環境で機能する器官であるため、陸棲化に伴う大きな変化はなかったといえる。そのためか陸棲化に伴う脊椎動物の味蕾に関する進化の観点での報告はほとんど無いようである。
感覚系のまとめ
感覚機能は摂食行動、回避行動、繁殖行動など生存上重要であり、これらの機能の陸棲化は、肉鰭類が浅瀬や潟に進出し、曝気の機会が増えると共に始まったと考えられる。
陸棲化の前にすでに脊椎動物はcolor visionに適応した受光細胞や筋肉などの眼球運動系、水晶体といった要素を含めた基本的な目の構造を確立していた。そのため陸棲化に際して形態的、機能的に大きな変化を要しなかったので、比較的早く大気中で機能する目を進化させたであろう。しかし、焦点調節方式を変えるには時間を要したであろう。
平衡感覚機能と聴覚機能を持つ内耳は無顎類の時代に既に獲得されていたが、肉鰭類は底生化やその後の浅瀬や潟への進出に伴い摂食や呼吸に際して顎や鰓の運動を制御していた舌顎骨の機能が低下した。一方曝気の頻度が増えると共に微弱な大気中の音波振動を拡大して内耳の聴覚部に伝播する必要が生じた。そのために第1鰓孔の外表面に鼓膜を形成し、鰓孔を中耳に変え、舌顎骨を鐙骨に変え、鼓膜の振動を内耳の聴覚部に伝播する仕組みを創りあげる必要があった。大気中で機能する聴覚器官の獲得は水中生活では不可能であり、実際に大気に曝されてから進化が始まったが、鼓膜を獲得した石炭紀初期の段階でも可聴範囲はそれほど広くは無かったであろう。小骨からなる中耳の形成には時間を要した。
陸棲化に際して嗅覚器官をあらたに作り上げる必要はなく、鼻腔内部構造や内鼻孔位置といった比較的小さな改変で対応出来たと思われる。臭覚レセプターの水溶性の臭覚物質検出機能を揮発性の臭覚検出機能に変える必要があったが、臭覚レセプターの多様性が大きいことから、その進化の速度は速く、改変にそれほど時間を要しなかったと思われる。従って、臭覚の陸棲適応は比較的短時間で可能だったであろう。
陸棲化が可能になった石炭紀初期には、感覚系は相応の適応を遂げていた。しかし、目の焦点調節機能、中耳や内耳聴覚部の複雑な器官構造の獲得には時間を要したと思われる。恐らくデボン紀末から石炭紀初期の四肢類は聴覚よりも視覚や臭覚に頼ったと思われる。