 魚が陸上を歩くまで
魚が陸上を歩くまで
第3章―1 骨格系
魚類から四肢動物へと進化する過程で骨自体の強化と頭蓋骨や全身骨格の構造に大きな変化があった。その過程を考察する前に、関連する主な骨の名前や骨の起源、構成を概略する。
頭蓋骨を構成する骨
無顎類では軟骨からなる内骨格とほぼ全身を被う皮骨で構成されていたが、進化とともに皮骨部は減少している。硬骨魚類と四肢類は共通した骨格構造をいくつか持っているが、初期の魚類では頭蓋部、特に口蓋やその周辺は非常に複雑ないくつかの骨で構成されており、しかも皮骨由来の骨と軟骨(内骨格)由来の骨が混在して結合している。さらに、魚類から四肢類にいたる過程で、頭蓋の皮骨要素が減っている。そのため頭蓋の進化は非常に複雑であり、魚類と四肢類の間で頭蓋を構成する骨の相同性が明らかでないものもある(Koyabu et al. 2012)。そのため様々の骨で構成された頭蓋部の、魚類から現生陸生動物への進化の過程は必ずしも明確ではなく、現在も研究と議論が継続中である(Nieuwenhuys 2009)。
頭蓋骨は内臓頭蓋と神経頭蓋に大別される。内臓頭蓋は主に顎、鰓を含めた顔面を構成する骨であり、上顎骨、頬骨(きょうこつ)、口蓋骨、下顎骨、舌骨、耳小骨などからなる。脊椎動物の基部に位置するとされるヤツメウナギはカンブリア紀の510Mya頃に出現したとされる。この頃に神経堤の形成があり、神経堤に誘導された内臓頭蓋由来の複雑な骨格の形成が可能になった。頭蓋部を構成する内臓頭蓋由来の骨には、初期のヤツメウナギとcrown-vertebrates:冠脊椎動物の間に明確な相関関係が見られない。このことは、既にヤツメウナギからcrown-vertebratesへと進化する過程において神経堤由来の間充織の全体的な、あるいはかなりの部分の再配列があったことを予想させる。魚類を経て四肢類へと移行するころには骨化した内骨格と軟骨から成る内骨格が主となり、皮骨由来の骨は頭蓋と鎖骨だけになった。
神経頭蓋は主に脳や臭覚器、視覚器、聴覚器など感覚系を覆う骨であり、前頭骨、側頭骨、頭頂骨、後頭骨などからなる。
顎口類の頭部は頭骨と下顎骨からなる。頭骨の上部には、比較的平板ないくつかの骨で構成される皮骨性の頭蓋天井とよばれる骨があり、その下にある脳函を保護している。頭蓋天井の前方には眼窩があり、その前方に口吻がつながっている。口吻には外鼻孔があり、眼窩と口吻から下側に向かって頬骨がある。口吻と頬骨の下には上顎骨が結合している。頭蓋天井の後ろ側は下顎骨と関節している。
肉鰭類では脳函は前後二つに分かれ、蝶番でつながっている。後ろ側の脳函は脊椎に関節している。脳函には、脳と平衡器官や聴覚、視覚、嗅覚といった感覚器官につながる神経が収められている。脳函は軟骨由来であり、内骨格である。
顎の起源は鰓弓とされる。魚類では咽頭部の後方に鰓弓という上下対になっている骨格が5対あり、これで鰓孔を支えている。鰓弓の先頭の一組が上下対の顎骨へと変化したと考えられ、上側の顎骨は脳函に結合している。
上顎骨は先端部の前上顎骨と主上顎骨が融合しており、歯骨を持つ。下顎骨は口蓋にある蝶番の機能を持つ方形骨で頭骨と関節しており、歯骨を持つ。鰓弓の下側は、顎の下の弾力性のある正中骨に接合しており、魚では呼吸により鰓腔が広がったり縮んだり出来るようになっている。上顎骨、下顎骨ともに皮骨由来である。
口蓋は脳函、口吻、頬骨によって形成されており、口中の天井となっている。頭骨の後ろは下顎の喉に向かって一連の皮骨からなる、鰓蓋咽頭骨系(さいがいいんとうこつ系)と呼ばれるいくつかの骨があり、鰓腔(えらこう)を保護している。
口蓋部はいくつかの舌弓(ぜつきゅう)や方形骨、関節骨、舌顎骨、翼状骨などで構成されているが、舌顎骨は特に重要な機能を持っている。舌顎骨は脳函後部にある耳殻に関節し、さらにいくつかの骨を介して方形骨につながっており、これにより口蓋の後部を支えている。従って舌顎骨は、呼吸や摂食に際しての口蓋と鰓弓の運動を制御している。四肢動物では口蓋に内鼻孔と呼ばれる外鼻孔とつながった孔があり、気道を経て肺へとつながっている。
全身骨格の構造
四肢類の体骨格は鎖骨以外は内骨格で構成されており、条鰭類の体骨格も内骨格で構成されているが、鰭条が皮骨由来として残っている。体幹部を支える中軸骨格はいくつかの椎骨からなり、魚類では対鰭以外の背鰭、尾鰭、尻鰭の骨も中軸骨格に含まれる。椎骨は、魚類では椎体の背側に神経棘が、腹側に血管棘が形成されている。神経棘や血管棘は部位によりサイズは異なるが、椎体の形状は後頭部から尾部にかけて似ている。
四肢類では椎骨は背側から腹側に向けて、脊髄を保護する神経弓、脊索を囲む椎体、および血管弓で構成された一つの単位であり、この椎骨が互いにいくつか関節して脊柱を形成している。胸部の椎骨には神経弓と椎体に肋骨が結合している。四肢類では、四肢の形成などにより椎骨の機能が部位ごとに異なってきたことを反映して、頸部、胸部、腰部で椎骨形状の違いが大きい(Mallo et al. 2009)。
付属肢骨格は、魚類では胸鰭と腹鰭を構成する一連の骨とこれら鰭と体をつなぐ肩帯(胸帯とも)と腰帯からなる。肩帯は小さな肩甲烏口骨や鎖骨等からなる。肩甲烏口骨には肩関節があり、胸鰭と関節している。鎖骨は皮骨由来であり、頭骨後部と結合している。軟骨魚の胸鰭は肩帯の近位から遠位方向に3本の柱脚骨(propterygium, mesopterygium, metapterygium)で構成されているが、条鰭類の鰭ではmetapterygiumが消失し2本の柱脚骨になっている( Zhu & Yu 2009)(Ahn & Ho 2008)(91 Cole 2007)。腰帯は内骨格由来の骨であり、魚類では体壁の内側に他の骨と結合することなく浮いた状態で固定されている。
四肢類の付属肢骨格は四肢骨と肩帯、腰帯からなる。初期の肉鰭類の鰭は軟骨魚類と同様3本の柱脚骨を持っているが、crown肉鰭類の段階ではmetapterygiumのみが残り、これが四肢類の上腕骨や大腿骨へと進化している。前肢では肩帯は大きな肩甲烏口骨や鎖骨等からなり、1本になった柱脚骨は肩関節で肩甲烏口骨に関節している。腰帯は魚類のそれよりかなり大きく、構造も異なっている。左右の腰帯が腹側に伸びて結合し、骨盤を形成している。背側は腰椎と尾骨の間の仙骨を介して脊柱と結合している。後肢の骨の構成は前肢のそれと似ており、1本の柱脚骨が骨盤に関節している。また、四肢の遠位部は指骨を含めた多数の自脚骨で構成されている。
骨の起源
骨の由来に関しては諸説がある。主に現生の原始的な脊椎動物や無脊椎動物の生理学に基づき、浸透へのバリア機能のため、必須ミネラルイオンの保存のため、産生された廃棄物に対する緩衝のため、産生廃棄物の処分のため等多くの仮説が提案されてきた。また、化石の詳細解析からセンサー機能向上のため、あるいは防御のためという仮説も出されている。特に骨が必須ミネラルイオンを保存する機能を持っていたために、淡水棲息や陸生化が可能になったという必須ミネラルイオンの保存仮説や脊椎動物出現時に食物連鎖のトップにいた海サソリからの防御のための外骨格甲皮形成仮説は有名である。
カルシウムを燐酸カルシウムに変換する機能は、多細胞化する段階で獲得していたとされる。この機能により脊椎動物の系統でもかなり初期の段階で過剰カルシウムを燐酸カルシウムの形にして貯蔵することが出来るようになっていた(Kawasaki & Weiss 2006)。カルシウムの小腸からの吸収にはヴィタミンDの活性化が必要であり、ヴィタミンDの活性化には太陽光が必要である。従って700Myaからカンブリア紀にかけてオゾン層が形成され、浅海に棲むことが出来るようになったことでカルシウムの吸収量が増え、骨の骨化が加速されたであろう(Young 2008)。遊泳能力を高めるための筋肉の支柱として強化した背骨は、魚類が淡水域に進出するときにはカルシウム不足を補うために機能した。また、汽水域でしばしば大気に体を曝した魚類は重力への対処のために各種骨格が強化された。
燐酸カルシウムの骨を持った最初の脊椎動物はコノドントと考えられる。コノドントは全体的な体型はヌタウナギやヤツメウナギに似ている。口内に象牙質の上をエナメル様の組織でカバーされた歯からなる複雑な歯列を形成している。最初にミネラル化した骨が形成されたのは、コノドントのfilter-feedingのための歯であったことから、骨のそもそもの起源は摂食用であり、その骨化のメカニズムを防御のための皮骨形成に適用したのが甲皮類の外骨格であるとされる(Donoghue & Sansom 2002)(Donoghue et al. 2006)。しかし、皮骨と内骨格では骨の形成過程が異なっている。内骨格は軟骨性骨化(endochondral ossification)によって作られる。即ち、最初軟骨として形成され、それが骨組織で置き換わる。一方皮骨は膜性骨化(membranous ossification)によって直接に骨として形成される。
従来歯は皮骨由来と考えられていたが(Donoghue et al. 2000)、コノドントの表皮に硬化組織は見られず、従って、コノドントの歯は皮骨由来ではないとされた(Janvier 2007)。しかし、内胚葉も歯の形成に関与していることが明らかになり(Soukup et al. 2008)、コノドントの歯は構造的にやや異なるところもあるが、脊椎動物の歯と相同な器官と考えられているようである。
近年の分子生物学的な手法も含めて、現生のヌタウナギ(Hagfishes)やヤツメウナギ(Lampreys)における骨形成の詳細が調べられ、一連の骨の進化に関与する遺伝子が明らかにされてきている(Hecht et al. 2008)(Wada 2010)(Zhang & Cohn 2008)(Zhang et al. 2009)(Zhang & Cohn 2006)。軟骨や骨形成に必須のfibrillar collagen に関連する遺伝子RunxとSoxEの共発現は頭索類において既に機能しており、これらの遺伝子はその後の遺伝子重複を経て脊椎動物の軟骨や骨の形成を可能にした。軟骨はdomain shuffling により、骨はtandem duplicationにより、新たな遺伝子を生み出すことで形成が可能になった。特に後者の遺伝子重複によりカルシウムと燐を制御するSCPP: secretory calcium-binding phosphoprotein geneが出来て、軟骨の骨化(ミネラル化)が可能になったとされる(Donoghue et al. 2006)。なお、これら遺伝子重複に先立って頭索類から円口類へと、filter feeding からactive predation へと摂食方式が変化する過程で外胚葉神経堤の形成があったとする説(Hecht et al. 2008)(Yu et al. 2008)と、遺伝子重複が外胚葉神経堤形成に必要であったとする説がある(McCauley 2008)。
ヌタウナギとヤツメウナギは咽頭部に内臓頭蓋由来と考えられる軟骨を持つが骨化していず、主成分は非コラーゲンのたんぱく質である。ヌタウナギ発生時の分子解析では硬節形成に必要とされるPax1とPax9の発現が見られないし、Pax1の活性化を司るNogginやSHHも無い。現生種のヌタウナギでは硬節は見られないが、中胚葉節が皮膚節と筋節に分化したとされる。一方ヤツメウナギでは筋節や硬節の形成が見られ、体軸性内骨格を形成しており、また尾鰭も持っている。従って、出現当初における頭索類は体軸性内骨格の基になる硬節が無く、もっとも早い骨のカルシウム化を獲得したのはヤツメウナギの段階と考えられる(Langille & Hall 1993)。なお、現生種よりも古生代種の方が骨化が進んでいるとされるが、現生種と古生代種の間の解剖学的な相関に関しては議論が継続中である。
魚類の骨格の進化
欠甲類:Anaspidsは尾鰭の形状や単鼻孔という点ではヤツメウナギに似ているが、骨化した多数の小さな鱗で覆われ、対鰭を持ち、背腹方向に動く下顎板を持っている。欠甲類の段階では小さくて貧弱な鱗を持っていた程度であったが、やや遅れて出現したとされる甲皮類:Ostracoderm:は充分に発達したarmorを持っており、表皮に硬質の外骨格を獲得した最初の脊椎動物とされる。従って皮骨は急激な進化で生じたと考えられる。こういった急激な皮骨の出現が可能だったのは、脊椎動物出現の初期には内胚葉と外胚葉の間の区別がなかったためであろうとされている(Smith & Cates 2001, 1998)。最近の有尾両生類の発生を詳細に観察した実験はこれを支持していると考えられる(Soukup et al. 2008)。
甲皮類の多くは頭部が融合した何枚かの大きな板状の外骨格でカバーされており、胴体部と尾部が皮骨性鱗で覆われており、対となっている胸鰭は顎口類の対鰭と相同と考えられる。頭部骨格にはカルシウム化が見られるが(Zhu & Janivier 1998)、尾鰭には内骨格は形成されていない。甲皮類の中には皮骨にエナメロイド層のある歯状突起を持つものが出現している。体軸性内骨格はヤツメウナギと大差の無いレベルであり、骨化は見られない。なお、頭部に板状の外骨格が無く軟骨魚類と同様頭部も小さな鱗で覆われている腔鱗類 :Thelodontsも対の胸鰭を持つが、顎口類の対鰭と相同かどうかは不明である。
甲皮類では全身を外骨格が被っていたが、その後魚類から四肢類へと移行する過程で外骨格は減少している。一方軟骨性であった背骨などが内骨格化している。燐酸カルシウムへの変換・蓄積機能を持っていたことにより、軟骨を燐酸カルシウムで置き換えることが出来たと考えられる。体骨格として燐酸カルシウムの骨を持った最も初期の脊椎動物は、無顎類のOsteostracansとされる。外皮は燐酸カルシウムの皮骨で覆われ、内骨格は軟骨で形成されていた。オーストラリア中央部の480Myaの地層から発見されたアランダスピスも目周辺に内骨格と思われる骨を持っていたが、燐酸カルシウムではなかったようである(Donoghue & Sansom 2002)。
甲皮類の1種であるガレアスピス化石の頭蓋を詳細に調べた最近の報告によると(Gai et al. 2011)、円口類や骨甲類といった他の無顎類とは異なり、1対の鼻嚢を持ち下垂管が口蓋に向かって開いている。この構造は顎口類と共通しており、顎口類の祖先系とされる無顎甲皮類の段階で既に顎の原型を獲得していたことがうかがえる。
鰓弓は、咽頭部の左右に、中央部で後ろ側に凸状に折れ曲がった形で対を作っている。その左右一組の鰓弓の折れ曲がった部分が関節でつながれて顎になったとされる。また顎補強のために、顎を形成した鰓弓のすぐ後ろの左右一組の鰓弓も変化している。即ち、この左右一組の上半分は頭骨と上顎とを結びつける舌顎骨となり、下半分は下顎を強化する舌骨になった。顎は皮骨由来と内臓頭蓋由来の骨で構成されていることになる。
上顎骨と下顎骨を関節する部分が実際にどのように進化してきたのかは明らかでない。そのため無顎類ヤツメウナギの咽頭部におけるいくつかの遺伝子の発生時における発現状況を、顎口類の顎周辺における遺伝子発現状況と比較した(Cerny et al. 2010)。その結果ヤツメウナギと顎口類では咽頭部の背腹軸を決める遺伝子Msx、Hand、Dlxの発現パターンが似ていることが分かった。一方、顎口類の第1鰓弓に見られるBapxとGdf5/6/7遺伝子はヤツメウナギでは見られなかった。このことから背腹軸を決める遺伝子プログラムMsx、Hand、Dlxに新たに顎関節部のプログラムBapxとGdf5/6/7が組み込まれることで顎口類の顎部が形成されたと考えられる。
一対の鰓弓から顎を獲得した最初の脊椎動物は板皮類:Placodermsである(Zalc & Colman 2000)。板皮類も甲皮類と同様に頭部はいくつかの板状皮骨が一体型に融合した頑丈な皮骨板となっており、また体部と尾部は皮骨鱗で覆われている。皮骨表面には、pore-canal systemsが見られる。内骨格は軟骨:perichondral bonesで構成されている。
なお、獲得された顎の構造は特に板皮類と肉鰭類では安定したものではなく、デボン紀を通じて変遷を重ねていたとされる。一方、軟骨魚類と条鰭類の顎の構造はデボン紀を通じて安定しており、獲得時の形状をほぼ保存しており、大きな変化は無かったとされる(Anderson et al. 2011)。そして顎の構造が不安定であった板皮類はデボン紀末に絶滅しており、肉鰭類もデボン紀末に大幅に種数を減らしている(Sallan & Coates 2010)。顎構造が安定であった軟骨魚類と条鰭類はデボン紀末の大絶滅事変の影響をほとんど受けず、むしろ種数を増やしている。即ち、獲得した顎の構造は急激に変化する生態系への適応を決める重要な因子であったようだ(Sallan & Coates 2010)。
軟骨魚類は頭部を含め体全体が小さい鱗で覆われており、鱗には歯と同様の構造を持つ歯様突起が形成されている。鱗以外の骨は内骨格性の軟骨であり、この軟骨の主成分は脊椎動物の軟骨と同様コラーゲンである。顎には歯を有するが、歯の化石は鱗の化石よりもかなり後の地層から出土しており、初期の軟骨魚類ではまだ歯を持っていなかった可能性がある(Miller et al. 2003)。
棘魚類:Acanthodiansの頭部はいくつかの外骨格板でカバーされ、胴体部と尾部は菱形鱗で覆われている。外骨格はエナメロイド表面層を保持していない。内骨格は軟骨:perichondral bonesで構成されているが、板皮類や軟骨魚類の軟骨よりも骨化が進んでいる。5個の鰓を持ち、鰓が皮骨性のカバーで覆われている種もある。初期条鰭類と似た形状の脳函は骨化した軟骨で出来ているが、完全な骨化はしていない。歯を持つ系統と歯を持たない系統がある。
初期硬骨魚類の頭蓋はいくつかの骨からなるが、骨間の結合はゆるい。内骨格は、前駆軟骨と骨化していない軟骨で構成されている。顎は可動性が高く、口を大きく開くことが可能であり、餌を丸呑みしていたことがうかがえる。また、上下顎には捕獲効率を高めるための歯がある。鰓弓:Gill arches は他の顎口魚類に比べると特に高度に進化している。肩帯は鎖骨と肩甲骨、烏口骨など幾つかの骨で構成され、これらは胸鰭の筋肉を支持する機能を持っている。
条鰭類は背鰭が一つであり、下顎と頬骨にはsensory-linesを持つ。口の開口は大きく、上下の顎には鋭い歯が発達しており、丸呑み式の摂食をおこなっていた。脳函の骨化は進んでいるが、体軸骨の骨化は初期条鰭類ではあまり進んでいない。鱗間の結合は棘魚類よりも強く、体部の剛性を高め遊泳力の向上が図られている。鱗の表面は金属光沢のガノイン:ganoine層を持ち、鱗形状は菱形が多い。後の条鰭類ではガノイン層が消失し、鱗はrounded shapesとなっている。
肉鰭類の脳函は蝶番で繋がれ前後に分かれている。多くは目が大きく、単一の外鼻孔を持つオステオレピス類:osteolepiforms 以外は対となった外鼻孔を持つ。総鰭類:Crossopterygiansではコズミン:cosmine型の鱗と皮骨にpore and canal systemが見られる。鱗同士のオーバーラップが大きく、板皮類や棘魚類よりも鱗間のつながりが強い。これにより体部の剛性が強化され、遊泳力が向上し、より活動的な行動が可能になったと考えられる。分岐直後の肉鰭類は祖先である初期硬骨魚類の遊泳性を引き継ぎ、広い生息環境に適応・放散したが、それを反映して鱗の形状も菱形や丸型など多様である。さらに肉鰭類の一部には底生へと移行した種もいた。丈夫な鱗を持つ群と軽量化された鱗を持つ群があり、前者が底生型、後者が遊泳型に対応していると考えられる。
魚類の内骨格、特に脊椎は体を支えるのではなく、筋肉の付着点の役割を持つ。急激で活発な運動のためには、付着点としての脊椎の強化が必要である。初期硬骨魚類から肉鰭類にかけて、脊椎を骨化することにより遊泳機能を強化してきた。底生、待伏せ型になると獲物を瞬発的に捕獲するために特に尾鰭部の骨化が強化されている。
肉鰭類の椎骨の形状は最初は魚類の椎骨と似ているが、底生化からさらに潟などの潮汐域への進出が進むと共に魚類様椎骨から椎骨同士のかみ合わせを強化した四肢様椎骨へと椎骨の形状を変えた。また、部位によって異なった椎骨形状をとるようになる。こういった椎骨の変化と同期して最初に胸鰭が、それに遅れて腹鰭が四肢へと変化した。
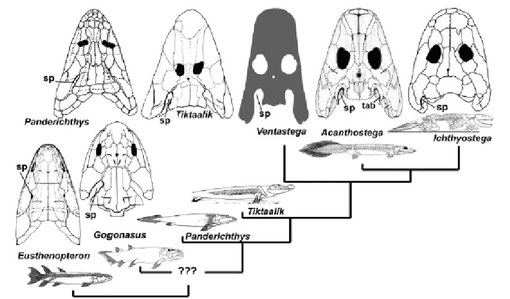
上述の骨格形成における進化の各ステップと系統の関係をまとめて図1に示す(Donoghue & Sansom 2002)。骨においては11段階の進化を経て肉鰭類に到っている。

大文字A~Kは骨格系に起こった顕著な事象を示す分岐点に対応。A:最初期の骨格形成、B: neurocranium の起源、C: 軸性内骨格における神経系の起源 D: ミネラル化骨、歯性骨の起源、E: 皮骨の起源、皮骨鱗の形成、F: 軟骨、ミネラル化内骨格の起源、G: 付属肢内骨骨格の起源、H: ミネラル化した軸性内骨格の起源、歯の起源、I: 鰓弓と歯の形成、J: neurocranium の骨化、K: 内骨格の骨化 From Donoghue & Sansom 2002
上陸前後における骨の変化
肉鰭類は420Myaには既に出現していたとされるが、陸棲化に結びつく変化は395Mya頃棲息していたKenichtysが内鼻孔を持ったあたりから始まったと考えられる(Zhu & Ahlberg 2004)。しかし、この内鼻孔の獲得は陸棲化と関係していたのではなく、底生への適応だったと考えられる。即ち、底生になり、特に待伏せ型の摂食方式になると鼻孔内の水流が保持できなくなる。鼻孔内の水流を得るために後外鼻孔を口蓋内に移し、鰓運動によって前外鼻孔から水を入れ内鼻孔から口中を経由して鰓から排出する経路を作ることで、嗅覚器官としての機能を保持した。やや扁平な頭骨、顎の歯骨縁には多数の小さな歯が見られる。こういった特徴からKenichthysは底生で、摂食方式は丸呑み方式であったと考えられる。
その後385~380Mya 頃棲息していたユーステノプテロン:EusthenopteronからTiktaalikあたりにかけて鰭に四肢的な特徴が顕著になっている。ユーステノプテロンにおいて2分割されていた脳函が一体化し始め、腰帯にも四肢類的特徴が生じ始めた(Chang & Yu 1997)。脳函の一体化は捕食方式が丸のみ型から噛み付き型に変化したことを(Lu & Zhu 2010)、腰帯の四肢化は泳ぐための鰭から這うための鰭へと変化したことを反映している。また、顎の歯骨の縁部には小さいが鋭い歯が一列に並んでいる。鰭における四肢的な特徴の出現は、ユーステノプテロンの頃に底生からさらに潮汐域や潟、三角州などの草木が茂った、あるいは泥濘の浅水域に進出したことにより移動方式が変化したためと考えられる(Clack 2006)。最も遠位の指骨を除くと肢を構成する基本の骨格はユーステノプテロンにおいて既に備わっている。また、通気孔が頭部後方に在り、これは泥濘等浅瀬を活動域にしていたことを示している。Kenichthysと同様に待伏せ型の丸呑み方式の摂食を行っていたとされるが、むしろ追跡型の噛付き方式で捕獲する様式に変わりつつある段階と思われる。
しかし、ユーステノプテロンは、頭部の形、目の位置、体型などから完全な浅瀬棲息ではなく、まだ底生が主であったと思われる。あるいは淡水域の環境、即ち海沿いの潟のような広範な浅瀬ではなく、狭い浅瀬にすぐ隣接して深みの水域があるような環境への適応だった可能性もある。
ユーステノプテロンからアカントステガやイクチオステガに到る系統は、四肢様魚類:Tetrapodmorhansと呼ばれる。ユーステノプテロンからアカントステガやイクチオステガに到る変化は、陸棲化への適応というよりは、浅い水域への適応と考えられる。四肢様魚類から初期四肢類までの頭部と体形の変遷の概略を図2に示す。ユーステノプテロンからアカントステガ・イクチオステガへと進むと共に頭骨が扁平化し、目が側面から頭頂部に移動し、胸鰭と腹鰭が四肢化し、背鰭が縮小・消失している。捕食方式は丸呑み型から噛付き型へ、呼吸方式は鰓機能を残しつつ肺機能の向上へ、移動方式は浅瀬を這う方式への適応変化と見られる(Clack 2006)。
頭骨の扁平化や目の背側への移動は、現生ワニ類のように浅い水域での摂食行動に適応したものであろう。顎や顎関節、歯は獲物の補足力が強化される方向に進化している。大きな開口を可能にしていた2分割型の脳函と顎関節を構成する舌骨弓:hyoid archは、大きく口を開く必要性が減ったことにより顕著な変化が見られる。摂食行動の変化に伴い舌骨弓は役割が低減し、縮小している。これにより頭部後方に空間的な余裕が出来、頸の形成と肩帯の大型化が可能になった。肩帯の大型化により胸鰭の肢的機能の向上が促された。また、魚類の舌顎骨:Hyomandibularは呼吸や摂食に関与する骨格系の動きを制御する重要な機能を持っていたが、四肢化に伴い舌顎骨が関節する舌顎骨関節骨が消失した。そのため顎関節部を形成していた舌顎骨はその機能を失った。Acanthostegaの段階になると舌顎骨は蝸牛窓近くに位置し、聴音機能を持ち始めた可能性がある(Clack 2003)。舌顎骨はその後聴覚に必要な鐙骨へと変化していくが、大気中で機能する聴覚になるまでの過程はかなりゆっくりしたものであった(ジンマー)。石炭紀の四肢類では舌顎骨は顎部における機能を失い、聴覚専用の微小骨となる。哺乳類では鐙骨に、爬虫類では耳小柱へと進化していく。
図はスケールを合わせていない。sp:通気孔 squ:側頭鱗 From Clack 2006
また、最初に胸鰭が、潟などの泥濘地において体を左右にくねらせて移動するための支点となったり、草木類をかき分けて進むために肢的な機能を持つようになり、さらに腹鰭の構造も追尾して骨格形状が変化・強化している。肢への移行は、胸鰭の方が腹鰭よりも先行したことは確かであるが、鰭から肢への変遷は多様である。最初は、体の屈曲による推進運動の段階、つまり地面への固着部分として腹鰭があまり重要でなかった段階を経て、パンデリクティスからアカントステガまでの間に後肢を使った推進運動が出現したようである。移動方式の変化に伴い、正中鰭のうち最初に背鰭の役割が低減し消失する。尾鰭の骨化は進んだが、水中における推進に必要な鰭先端部分は縮小する傾向を示している。
鰭形状や体型からパンデリクティスも水中で餌を摂っていたと考えられているが、パンデリクティスと四肢類の顎の構造は似ている。従ってパンデリクティスでは摂食方式が噛み付き方式に変ったが、咀嚼せず丸呑みするだけであったと思われる。頭蓋の縫合線の詳細形状から、水棲の丸呑み方式から陸棲の噛み砕き方式に変わったのはアカントステガ以降であるようだ(Markey & Marchall 2007)。
Tiktaalikの胸鰭や肩帯は肢化がさらに進み、大気中に露出した前半身の体重を胸鰭で支えることが出来たとされる(Daeschler et al. 2006)(Downs et al. 2008)。柱脚骨は既に1本になっており、胸鰭の運動機能が向上したと思われる。浅瀬において胸鰭を使った高速の移動が可能だったと思われる。
曝気の機会と時間が増えるに伴い付属肢を含め大部分の骨は実際に重力を感じる状況の中で進化したと考えられる。アカントステガやイクチオステガでは、水から出た背側の重さに耐えて這うために四肢が発達し、増大した腹部と水底の接触抵抗に対応するために移動の支点となる指が形成された。
底生の魚類は砂等の吸引から鰓を保護するために頭頂部に呼吸孔:spiraclarを持つものが多い。初期のシーラカンス類では見られないが、ハイギョの系統とされる413Mya頃出現したOnychodontや410Mya頃に出現したYoungolepis、さらには400Mya頃棲息していた条鰭類とされるLigulalepisには呼吸孔が見られる(Clack 2006)。
肉鰭類でも、頭頂部から口蓋に向かう呼吸孔が口蓋で開いており、頭頂部から口蓋への経路は舌顎骨や口蓋方形骨、脳函で囲まれた空間として形成されている。魚類段階にあった初期の肉鰭類の呼吸孔の経路は細くて曲がっているが、四肢類化とともに舌顎骨や口蓋方形骨が変形・変位し、四肢類様魚類段階のPanderichthysでは、呼吸孔の経路は広くまっすぐになっている(Brazeau & Ahlberg 2006)。
呼吸孔とその経路が水で満たされていたのか、空気で満たされていたのかに関しては明確ではない。初期硬骨魚の段階で肺を獲得していたのだから呼吸孔は通気孔であった可能性がある(Clack 2003)。しかし、鰓呼吸が主で、呼吸孔は底生における鰓保護のためであると考えられ、呼吸孔とその経路は水で満たされていた可能性が高いと思われる。アカントステガやイクチオステガは鰓蓋を持っており主には水棲であったが、餌の捕獲時などある程度の時間大気に晒されることに耐えられたとされる。現生両性魚類のトビハゼは、陸上では鰓呼吸と鰓の保護のために口中に海水を含んでいる。鰓が曝気に脆弱なことを考えると、初期両生類も大気に晒されて移動する時には、鰓蓋を閉じ、呼吸孔とその経路を水で満たし、鰓呼吸や鰓の保護をしていた可能性が高い。
大気中で呼吸をする四肢類の特徴とされる内鼻孔は肉鰭類Kenichthysにおいてすでに獲得されていたが、アカントステガ段階でも内鼻孔は口吻の先端近くにある。従って、呼吸のための空気を口から飲み込んでおり、鼻孔を呼吸には使っていなかった (クラック)。当時の両生類の扁平な頭骸骨と口吻は口から空気を飲み込む呼吸方式に適応した形状と考えられる。
アカントステガ段階では鼻孔が大気中での臭覚器官としても機能していた可能性はあるが、不明である。鼻孔が陸棲化に際して呼吸機能を持つようになったのはアカントステガよりはかなり後とされる。恐らく「ローマーの空隙」後に出現した頭部が扁平でない羊膜類が出現した頃に口を使った空気を飲み込む方式から鼻孔を使った呼吸へと移行したと考えられる。
陸生に必要な機能・構造は水中生活では獲得できないので、付属肢や頭蓋骨を含め大部分の骨は実際に重力を感じる状況の中で進化したと考えられる。曝気の機会と時間が増えるに伴い重力の影響を少なくするために背側の体組織を減らす方向への選択があったと予想される。この結果脊椎は魚類のような体中央ではなく、背側に偏在するようになった。これは背側の体幹筋肉の減少を、従って尾鰭を左右に振って移動する機能の低下をもたらしたであろう。図3に見るように、背骨の偏在はユーステノプテロンにおいて既に見られる。アカントステガでは、曝気に伴う加重から内臓を保護するために肋骨の発達が顕著である。
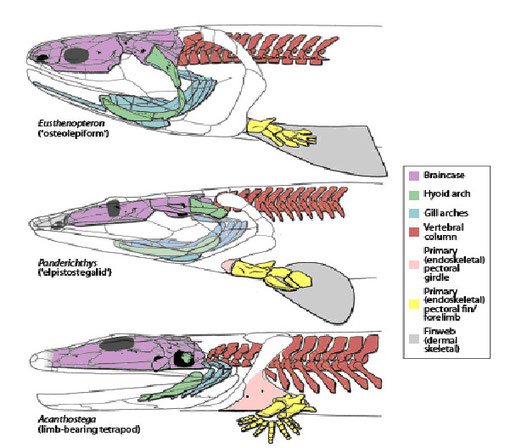
頭蓋、顎、鰓弓、背骨、胸鰭、肩帯の浅水域への適応に伴う変化を示す。
From Coates et al. 2008
パンデリクティスの胸鰭は上腕骨や尺骨、橈骨に対応する骨と指に似た遠位放射骨と鰭条骨を持っている。しかし、腰帯はユーステノプテロンのものほどには四肢類に似ていない(Boisvert et al. 2008) (Boisvert 2005)。ティクターリクの胸鰭も上腕骨や尺骨、橈骨に対応する骨や遠位放射骨を持つが、さらに肘関節と手首関節が加わり、また遠位放射骨も関節でつながっている(Yano & Tamura 2012)。胸筋も発達しており、前鰭により水面上に体を持ち上げることが出来たとされる(Shubin et al. 2004)。
鰭から四肢への変化には、指や仙骨のような新しい形態の生成を伴うと共に、移動運動を担う器官が胸部付属肢から腹部付属肢へと重点を移行したことも含まれていた。こうした変化はユーステノプテロンから初期四肢動物のイクチオステガやアカントステガに至る間に起こっているが(152 Boisvert et al. 2008)、腹鰭から後肢への変化を示す化石は少なく、化石によって変化のようすが直接明らかになっているわけではない。
表1にユーステノプテロン、パンデリクティス、チクターリク、アカントステガの特徴を一覧にまとめる(Ahlberg et al. 2008)(Ahlberg & Clack 2006)(Boisvert 2005)。
表1.陸棲化における特徴の変遷
|
種名、Mya |
頭蓋骨の特徴 |
全身の特徴 |
その他 |
|
ユーステノプテロン Eusthenopteron 385Mya 体長1.2m |
丸い頭蓋、短い吻部と長い脳函 後頭頂骨が長く、蝶番関節の痕跡 脳函は前部と後部の二分割 脳函の骨化は不十分 吻部はモザイク状骨で形成 目は側面,眼窩は小さい 外鼻孔は小さく,吻の先端位置 骨化した鼻殻、頭蓋 鰓蓋咽頭骨あり 多数の細かい歯 |
全身が皮骨性鱗で覆われている 鱗のエナメル/エナメロイド、象牙質の消失→軽量化 成長とともに骨化部分増加 骨化は後部ほど顕著、胸鰭、腹鰭に骨化見られず:石炭紀両生類と逆 頭骨に接続した複数の擬鎖骨が肩帯に存在 軟骨性肩甲烏口骨は3点で皮骨性擬鎖骨に関節 腰帯はパンデリクティスより四肢類的 上腕骨,橈骨,尺骨に対応する骨を持つ 大腿骨、脛骨、ヒ骨に対応する骨を持つ 対鰭の先端に扇状の皮骨性の鱗状鰭条が伸びて鰭を支えている 背鰭、尻鰭あり、尾鰭の端は皮骨性の鱗状鰭条が上から下まで縁取る |
河口~海生 植物が繁茂する底床に棲息 体を左右に振る魚類と同じ推進方式 待伏せ型 吸込み式摂食 |
|
パンデリクティス Panderichthys 378、385Mya 体長0.9~1.3m |
扁平頭蓋, 吻部の長さは中間, 吻部は大きな骨で形成 後頭頂骨が長い 頭頂骨に蝶番関節の痕跡無し 脳函は前部と後部の二分割 頭頂に鰓呼吸孔 目は背側,眼窩は大きい 鼻孔は吻の低い位置に形成 骨化した鼻殻 鰓蓋、咽頭骨の縮小 舌顎骨縮小するも顎機能あり 細長く鋭い歯 |
全身が皮骨性鱗で覆われている 頭骨に接続した一個の擬鎖骨が肩帯に存在 軟骨性肩甲烏口骨が拡大、全面で皮骨性擬鎖骨に関節 胸鰭は前肢に近い形状 前肢による頭部の支持は可能だが前進機能はほとんど持たず 腰帯に四肢類の特徴が見られる 後肢は推進のためのアンカー機能 対鰭の先を皮骨性の鱗状鰭条が縁取る 尾鰭の端は皮骨性の鱗状鰭条が上から下まで縁取る 背鰭、尻鰭無し 鰭先端に指は無いがティクターリクより四肢類的な鰭構造 |
海岸縁、干潟 浅瀬泥濘棲息 後肢を支点にし左右に振る推進方式 噛付き式摂食 |
|
ティクターリク Tiktaalik 375[380]Mya 体長1~2m |
扁平頭蓋,吻部は長い 頭蓋後部が短い 長い口吻を持ち、吻先端に鼻孔 脳函は二分割、頭皮に分割痕跡無い 目は背側、大きくてくっついた眼窩 口蓋構成骨はパンデリクティスに似る 口ポンプ機能の向上:鰓,肺呼吸 鰓呼吸孔 鰓孔と喉部を覆う骨が縮小 舌顎骨の縮小、再配置化 多数の小さくて鋭い歯 |
皮骨性鱗の減少 軟骨性肩甲烏口骨が拡大、全面で皮骨性擬鎖骨に関節し、両骨とも骨化進行 鰓蓋、外肩甲骨系消失→頸を持ち、可動 肋骨が発達,隣接同士で重なる 隣接肋骨同士の重なり顕著 肋骨の骨化は進むが、椎骨骨化は不十分 胸筋が発達 上腕骨,橈骨,尺骨に対応する骨を持ち、肩帯に関節 前肢の可動範囲はパンデリクティスより大、アカントステガより陸棲化 肘,手首関節を持つ 鰭には放射骨があり,先端に鱗状鰭条 |
熱帯の浅い水底、淡水の氾濫原の泥地に生息 四肢による上体支持可能 噛付き式摂食 |
|
アカントステガ Acanthostega 365Mya 体長0.6m |
扁平頭蓋を持ち、後頭頂骨が短い 吻部は大きな骨で形成、丸く長い 鼻孔は吻先端下部 頭頂骨に蝶番関節の痕跡無し 脳函の前部後部の区切り無し 目は背側,眼窩は大きい 鰓蓋と咽頭骨無いが、鰓骨格、鰓弓に動脈溝、後鰓板により鰓,肺呼吸 耳構造はパンデリクティスともイクチオステガとも異なる 内側に小さな歯列、外側に大きな歯と牙 |
腹部のみ長卵形皮骨性鱗 椎骨形状はどこでもほぼ同じで魚類に似る 頸の可動性はかなりある 軟骨性肩甲烏口骨が拡大、全面で皮骨性擬鎖骨に関節し、両骨とも骨化進行 橈骨は細長く、尺骨の2倍の長さ、尺骨は短く扁平で前脚は鰭形状 上肢の可動性はあるが、筋肉付着突起は無い 背骨に骨盤が関節し、大腿骨は上腕骨よりやや長い 尾は長い鰭条があり、鰭膜は背側から腹側まである 背鰭、尻鰭無し 8本指、手首関節は無く可動性は無い 鰓機能が主:内鰓呼吸か |
熱帯モンスーン 流速川底 四肢は体を支えられず 生息域はイクチオステガと異なる 水棲生活が主 噛付き式摂食 |
ほぼ同じ時期365Myaに異なる環境に棲息していたイクチオステガとアカントステガの全身骨格を図4に示す。両者とも腰帯は背骨に関節しており、強化された四肢を持っている。背骨の椎骨形状も部位によって異なっている。こういった特徴からかなり陸棲化が進んでいることがうかがえるが、しかしいずれも陸上歩行が常態ではなく、水棲を主としていたと考えられている(Clack et al. 2003)。また、最近の上腕骨の形状比較から、従来と異なり、イクチオステガの方がアカントステガより原初形態であると考えられている(Callier et al. 2009)。

a. イクチオステガ;頭蓋で隠れる最前部頸骨は灰色で示す。大きな前駆仙骨椎が見られる。従来の復元図に比べて、歯形状、指の方向が異なり、肩帯骨や腰帯骨がより大きく、尾がより短くなっている。前腕は、先端部環節の詳細が不明だが、強力なストローク運動可能性を示す。頸部と尾根元部の脊椎骨形状も不明。1:頸部 2:胸部 3:腰部 4:仙骨部。仙骨は灰色で示す。 c. アカントステガ;Coates の未発表データ、Coates (1996)とClack (2002) を元に作成。仙骨は灰色で示す。 スケール線は100ミリ
From Ahlberg et al. 2005
原初形態とされるイクチオステガの移動方式、即ち腹鰭、あるいは後肢があまり機能しない推進運動としては、体を背腹方向に屈曲させて移動する方式が考えられる。イクチオステガでは特に頑強な後肢と重畳した幅の広い肋骨が顕著である。重畳した肋骨や背骨、後肢の特殊な形状から、体部を左右に振る魚類型の、あるいは現生オオサンショウウオのように四肢を支点にして体全体を左右にくねらせる移動は出来なかったと考えられる。また、肩関節の3次元的動きを解析した最近の報告では、体を持ち上げた状態で肢を前後に動かすことが出来なかったとしている(Pierce et al. 2012)。これらのことから、イクチオステガは、多くの現生肉食哺乳類が獲物を追いかけるときやワニ類が獲物を捕獲するときのように、体部を背腹方向に振る移動方式を採っていたことが予想される(Callier et al. 2009)。イクチオステガの推進運動方式は後鰭、乃至は後肢をあまり使わない推進運動方式の名残であると思われ、後肢を使ったアカントステガの移動方式に到るまでに採用していた古い方式であった可能性がある。実際イクチオステガ以外でもいくつかの初期四肢類において重畳した肋骨が報告されている(Ahlberg et al. 2005)。
しかし、体型と運動様式の大きな違いは棲息していた生態系と生活様式の違いによる可能性もある。イクチオステガは非常に浅い潟のような泥濘水域に生息し、獲物を追うとき後肢は体が後ろに戻らないように機能した。一方、尾鰭に鰭部分を多く残すアカントステガの方はより水深のある浅水域に棲息し、体全体を左右にくねらせ、四肢を使って移動していたことも考えられる。
これら初期四肢類はおおむね大型であり、幅の広い扁平な頭骨と側線を持ち、指の数もアカントステガやイクチオステガ、チュレルペトンのように6~8本とまだ定まったものではなかった。デボン紀に40~50種ほどの四肢類の化石が出ているが、指を持つのはアカントステガ、イクチオステガ、チューレルペトンの3種のみである。アカントステガとイクチオステガは浅瀬の水環境における移動を行っていたが、チュレルペトンは陸上歩行が可能と考えられている(Long & Gordon 2004)。また、アカントステガやイクチオステガは肺のほかに鰓も保持していたが、チュレルペトンは鰓を持たず完全な肺呼吸に移行していたようである。
アカントステガ、イクチオステガ、チュレルペトン等の両生類が出現した後ユーラメリカ大陸とゴンドワナ大陸間での四肢類の往来があり、デボン紀末には地域特有性が薄れ始めた。しかしその時期に二回目の大絶滅が起こり(Clement et al. 2004)、これら初期の両生類の多くは生き残れなかった(Carrol 2002)。デボン紀末から石炭紀初期の345Myaまでのおよそ15My間は「ローマーの空隙」といわれる脊椎動物の化石がほとんど無い時代が続く。
現時点では、この空隙を埋める化石としてスコットランドの350Myaの地層から出たペデルペス:Pederpesとそれより若い346Myaの地層からのワトケエリア:Whatcheeriaの2種が知られているのみである。どちらも初期四肢類の扁平な頭部と異なり高さのある大きな頭部を持ち、似たような体形をしている。また、肋骨はイクチオステガのように幅が広く、脊椎骨もイクチオステガに似ており、類似の摂食行動をしていたと思われる。四肢の形は初期四肢類に似ているが、5指でありつま先が前方を向いているので、より陸上歩行に適した形になっている。後肢が体を支持する機能を持つのは前肢より遅れたが、「ローマーの空隙」後には前後の四肢が体部を支持出来る構造となり、地面をとらえる機能を持つ指を形成したことにより完全な陸上歩行が可能になった。しかし、側線系があり太い尾を持つことから、初期四肢類よりも陸棲化してはいるが依然として水棲主体であった。Pederpesは淡水域、Whatcheeriaは海性の浅瀬に生息していたとされる。
上陸後の四肢類の化石によると、上腕骨、大腿骨ともに陸棲化の進行に伴って細く、長くなり、陸上歩行に適した形になっていることを示している。また上腕骨の近位の関節面が変化し、橈骨や尺骨にすばやく力を加えることが出来るようになっている。同時に平行して肩関節も変化し、上腕の可動範囲が限定され、効率よい歩行が出来るように変化している。こういった変化は重力下での移動を必要とした上陸後に起こっている。
初期四肢類から羊膜類の分岐に到るさまざまの分岐系統図が提案されている (Laurin 2004) (Berman 2000). (Benton 1990)(Ruta et al. 2003)(Coates et al. 2008)。デボン紀末から石炭紀初期は急激な分岐・多様化の時期であり、相互に特徴がモザイク状になっているため、これらの系統図では細部はもとよりかなり大きな分類でも必ずしも一致はしていない。しかし、化石記録は少ないが「ローマーの空隙」の時期に既に多様化が始まっていた点では一致している。「ローマーの空隙」の時期に水棲種を主とするコロステウス網:Colosteudaeや多様な種を生み出した分椎目:Temnospondyliや炭竜目:Anthracosauria、現生両生類につながる両生類や爬虫類形目などが分岐し、さらには現生哺乳類の祖先系とされる単弓類:Synapsidaの系統も出現していた。
こういった準備期間があったため、四肢類は「ローマーの空隙」が終わった直後の石炭紀初期のヴィゼアン期、335Myaには急激に放散した。迷歯亜綱:Labyrinthodontiaの主な3つのグループとされる分椎目と炭竜目、及び空椎亜網:Lepospontyliのおのおので多様化している。新たに出現した四肢類は、いずれも初期は小型でほとんどが5指を持っていた。これら新四肢類の多くは水棲であるが、陸棲種も増加し始めた(Clack 2001, 2002)。多くは氾濫原や河口域、河川といった淡水環境に棲息する両生類であったとされるが、海生の四肢類も存在していた(Garcia et al. 2006)。さらには欠脚類もこの頃既に棲息していた。レティスクスは四肢を失っているが、鎖骨や脊椎骨、肋骨などに四肢類の特徴を持っている。即ち、ヴィゼアン世中期336Mya頃には欠脚類のように特殊化した四肢類が出現するほど多様化が進んでいたことになる。
従って、「ローマーの空隙」は、アカントステガやイクチオステガなどほぼ絶滅した初期四肢類のニッチを占めるべく種々の環境で様々の試みが為されていた新四肢類の揺籃の時代であった(Carrol 2002)。
分椎目は初期四肢類のイクチオステガの系統を引き継いでいると考えられ、頭骨も体部もやや扁平であり、現生の有尾目両生類に似た体形をしたものが多い。口蓋には現生両生類と同様空隙がある。口蓋空隙の外側を伸縮性の皮膚がおおっていて、呼吸や摂食のための機能を果たしていたのであろう。顎部の関節は可動性が無いので摂食は噛み付き方式だったと思われる。歯は迷路歯である。背骨の椎心は間椎心と側椎心の2個で構成されている。ユーステノプテロンの時期に椎心に新たに側椎心が加わり、間椎心と側椎心の2個で構成され、イクチオステガでは側椎心の比率が大きくなっている。しかし、分椎目では側椎心が縮小しており、ペルム紀の水棲種では再び魚類のように1個の間椎心だけになっている。椎骨のこういった違いは、陸棲と水棲に関係があると思われるが、あまりはっきりしていない。椎骨同士の結合の強さと関係があるのであろう。初期には皮骨性の鱗を持つ種類もあったが、後には鱗のない表皮になっている。前肢の指は4本、後肢の指は5本のものが多い。現生両生類の祖先とする説が一般的である(クラック)。
初期の分椎目の典型はバラネルペトン:Balanerpetonとデンドレルペトンであり、どちらも小型で陸棲である。従って分椎目は本来は陸棲であったと思われるが、後には陸棲、半水棲、水棲と多様化した。陸棲種では、側線系と鰓弓が無く頬部の後方に耳切痕があり、ここに鼓膜があったとされる。鐙骨は小さくて細い棒状であり、既に音波伝播の機能を持っていたとされる。しかし、陸棲種はペルム紀には衰退・滅亡している。
一方、炭竜目は現生トカゲ類に似た体形をした種類が多く、陸棲、半水棲、水棲と多様化しているが、分椎目ほどには繁栄しなかったようで、化石は多くない。エンボロメリ亜目:Embolomeriが最も繁栄したグループである。頭部も体部も幅が狭くなり、口蓋に空隙が無いので、口から空気を飲み込むのではなく、肋骨を使い鼻から空気を吸込む肺呼吸が可能になっていたことをうかがわせる。顎関節はあるていど可動性があったとされる。頭頂部骨格構造は羊膜類の構造に似ている。イクチオステガに比べて側椎心の比率がより大きくなり、間椎心の方がより小さくなっている。間椎心の縮小と側椎心の拡大は羊膜類と共通した特徴である。椎骨のこのような変化は背骨同士の結合を強めるためと思われる。陸棲種の四肢は短いが頑強であり、陸上歩行が可能であった。歯は迷路歯であり、指は陸棲も水棲も前後肢とも5本である。炭竜目は分椎目よりも爬虫類に近いとされる。
空椎亜網は比較的小型の種類が多く、分布も分椎目や炭竜目より狭く、生態系に占める位置は低かったと思われる。椎心の形成過程は現生の有尾両生類に似ているとされる。歯は前2者と異なり、迷路歯ではない。空椎亜網の系統から現生の平滑両生類が生じたとされるが、議論が継続中である。
スコットランドの340Myaの地層から出土したウェストロシアーナ:Westlothianaは両生類と羊膜類の中間の特徴を示している。双弓亜網とされ、椎骨や側頭部、前頭骨や眼窩周辺部の骨は陸棲脊椎類の特徴を持っている。また耳切痕が無く、歯も迷路歯でないので、羊膜類とされた。しかし、足首には羊膜類の特徴である距骨と踵骨(しょうこつ)が無いので、両生類の段階とする見解もある(Laurin 2004)。
炭竜目の系統とされるシームリア:Seymouriaは両生類と羊膜類の中間の形態を持っている。体骨格の特に椎骨が初期爬虫類の特徴を示している。一方、頭骨はエンボロメリの特徴を持ち、耳切痕や鐙骨といった聴覚関連部は分椎類に近い。半乾燥地帯に生息していたと思われ、雌雄の別があり、雌とされる個体の尾椎形状から羊膜卵を産んでいた可能性が指摘された。しかし、最近では羊膜類よりは両生類に近いとされている。
羊膜類の顕著な特徴は、3個の足根骨が癒着し一体化して距骨になっていること、上腕骨が扁平でなくなっていること、間椎心が小さくなり、側椎心が主になっていること、肋骨が発達し腹部をほぼおおっていること、耳切痕が無く鐙骨は顎の関節部に組み込まれていること、などである。こういった特徴を備えた、明確に羊膜類と判断される最初の四肢類化石は315Myaに棲息していたヒロノムス:Hylonomusである。また310Mya頃棲息していた単弓類のアーケオシリス:Archaeothyris、さらに300Mya頃生息していた双弓類のペトロラコサウルス:Petrolacosaurusも明確な羊膜類である。
これら羊膜類の化石と棲息年代から、羊膜類は「ローマーの空隙」の間に、また現生哺乳類につながるとされる単弓類も「ローマーの空隙」の間に爬虫類形目の中から分岐していたと考えられている。また、現生爬虫類の祖先といわれる双弓類は「ローマーの空隙」が終わった直後に分岐したとされる。即ち、現生哺乳類の祖先と現生爬虫類の祖先とは羊膜類が出現したほとんど最初の段階から別々に進化してきたことになる(Coates et al. 2008) (Zhang et al. 2005)(Laurin 2004)。
まとめ
対鰭や顎の原型は無顎類の時代にすでに獲得されていた。シルル紀前半には遊泳性の初期硬骨魚類が出現し、シルル紀後期には初期硬骨魚類から遊泳性の肉鰭類が出現した。
肉鰭類は395Mya頃から底生化を進め、それに伴い内鼻孔を進化させ、頭頂部に呼吸孔を発達させた。さらに385Mya頃に潟や三角州といった潮汐域や草木の繁茂した浅い水域の環境へ進出、頭部の扁平化、椎骨形状の部位による異形化、肋骨の形成、付属肢の四肢化、指の形成、背鰭と尾鰭の縮小を進めた。その結果アカントステガ、イクチオステガ、チュレルペトンなどの四肢類が出現した。これら初期四肢類は浅い水域の環境に適応したものであり、完全な陸棲には到らなかったが、ある程度の陸棲適応が可能になった。しかし、デボン紀末の大絶滅事変により初期四肢類はほとんど子孫を残すことなく滅亡した。
石炭紀に入ると初期四肢類に代わり新たに小型の四肢類が出現した。これら新四肢類は完全な陸棲を果たした種類もいたが、水棲種も多かった。石炭紀初期の360~340Myaの間にこれら小型の四肢類はトカゲ様、ウナギ様など陸上や水域で多様化しはじめ、羊膜類も出現した。「ローマーの空隙」は脊椎動物陸棲化の揺籃期であった。石炭紀のヴィゼアン期になると完全に陸棲化した新四肢類は各所で適応・放散を始めた。