 魚が陸上を歩くまで
魚が陸上を歩くまで
第3章―2 歯
歯は、棲息時の食性や生活環境を知る手がかりであり、化石として残りやすいために古くから多くの研究があり、関連情報が多い。しかし、意外にも歯の由来や顎と歯の進化の過程は依然として明確ではなく、今でも議論が継続中である。
軟骨魚類の皮骨性の楯鱗は歯と似たような構造を持っており、相同と考えられた。即ち、軟骨魚類の鱗が歯の起源とされた。また、顎を獲得したのは板皮類が最初であり、その後棘魚類や軟骨魚類の段階で歯を獲得したとされていた。実際顎が無ければ歯の生える場所がないという常識的な考えからすれば当然の帰結であり、この説は長いこと信じられてきた。
現生種を含めて多くの脊椎動物の歯は通常3層構造をしている。表面は硬質のエナメル、又はエナメロイドで覆われており、その下に多孔質の象牙質があり、さらにその下に骨質層がある。初期軟骨魚類の歯の表面は単層のエナメロイド層で、一方初期肉鰭類の歯の表面はエナメル層で覆われている。歯の表面層はより硬くて丈夫な方向へ、即ち、多層化とエナメロイドからエナメルへと進化した傾向を示す。従来説に基づくと、分岐図上における歯と歯の表面構造の獲得時期は下図のようになる(Gillis & Donoghue 2007)。なお、棘魚類ではエナメロイド様のカバーが見られないとの報告もあり(Donoghue & Sansom 2002)、歯の表面構造は可変性のある組織であるようだ。

顎が形成された後に歯を獲得しているとする従来モデル。 EA:単層エナメロイド EB:3層エナメロイド EC:エナメル層
From Gillis & Donoghue 2007
一方、無顎類Anaspida等の初期脊椎動物の外皮には皮骨:dermal skeleton性の歯状突起:dentine scale coneが形成されており、歯状突起の3層構造が、歯の3層構造、即ち表面が硬質のエナメル、又はエナメロイド層、その下に象牙質層、さらにその下に骨質層を持つ構造に似ていることから、相同性への疑問はあるものの(Downs & Donoghue 2009)、これが歯の起源とする説が提案されている(Reif 1982)。顎が出来てそこに歯が生じたことは確かであるが、歯そのものの起源は顎とは独立であったとされる(McCollum & Sharpe 2001)(Smith 2003)。板皮類の顎に歯が無かったことから、顎を持ってもすぐに顎に歯が形成されたのではなく、外皮の皮骨性歯状突起が顎の形成後に前方から口の中に入り込んだとするoutside-in hypothesisが提案された(Donoghue & Sansom 2002)。
この説だと歯の前駆体というべき皮骨が既に無顎類のAnaspidsに見られる。しかし、Thelodontsの段階でもエナメルのような硬いカバーは形成されていず、薄い単層のエナメロイドで覆われた歯状突起であった(Huysseune et al. 2009)。そしてこの構造は前述した初期軟骨魚類の歯の構造に似ている。最初に顎に歯を持ったのは軟骨魚類や棘魚類であるが、初期の軟骨魚類の中には歯を持たない種類があった(Sansom et al. 2000)(Sansom et al. 1996)(Smith & Johanson 2003)。また、軟骨魚類の鱗は455Mya頃の地層から出土しているが、歯の化石出土は418Nyaとかなり遅れており、軟骨魚類は初期には歯を持っていなかった可能性もある。棘魚類では3目のうち歯を持っているのはIschnacanthids1目に過ぎない。こういった化石情報はoutside-in hypothesisを支持していると思われる。
さらに、この仮説は歯を形成する4つの組織、骨、象牙質、エナメロイド、エナメルのミネラル化に関与する遺伝子SCPP (secretory calcium-binding phosphoprotein) family の発現を中心に魚類、両生類、哺乳類で調べた結果からも支持されるとしている(Kawasaki & Weiss 2008, 2006)(Kawasaki et al. 2007, 2005, 2004)(Donoghue et al. 2006)。これら一連の遺伝子発現実験から得られた結果を図2に示す。
SCPP familyの最も近い共通祖先型は、神経組織形成に関与するSPRCL1である。SPRCL1は象牙質と骨の非コラーゲンたんぱく質形成に関与するSPRCLから生じた。SPARCL1の出現はヤツメウナギとの分岐後、軟骨魚との分岐前であり、脊椎動物の初期に起こった全遺伝子重複:whole gene duplication 2(WGD2、またはR2)と関連している。また、SCPPの出現は条鰭類と肉鰭類の分岐の前であり、SPRCL1に起こったタンデム型の遺伝子重複と関連している(Kawasaki et al. 2007)。その後SCPPは肉鰭類と条鰭類で独立に進化し、肉鰭類ではエナメル形成を可能にした。エナメルはエナメロイドの構成たんぱく質以外のたんぱく質を持つのでエナメロイドから誘導されたものであり、歯のミネラル化は骨・象牙質・エナメロイド・エナメルの順で進化した。
上述のoutside-in hypothesisでは、皮骨が歯の前駆体であったとしており、従って歯は外胚葉由来の器官として生じたことになる。
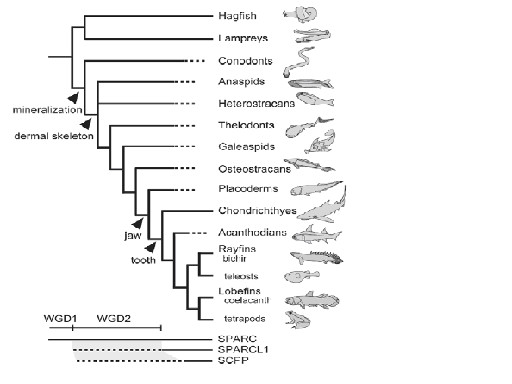
コノドント:conodonts:最も初期にミネラル化した骨を持った。最初に幹無顎類においてWGD1が機能し、その後幹顎口類においてWGD2が機能した。SPARCL1はWGD2によりSPARCから生じた。SPCCはSPARCL1由来であるが、現生種では硬骨魚類と四肢類にだけ見られる。最初のSPCCは条鰭類と肉鰭類が分岐する前に出現した。分岐図はDonoghue et al. (2006) に基づいている。 WGD:Whole Gene Duplication
From Kawasaki 2009
一方、口中の歯の起源に関してはinside-out hypothesisも提案されている。この仮説は咽頭部に形成されている渦型歯状器官(pheringal denticles) が歯の起源であるとする説である(Smith & Coates 1989)(Johanson & Smith 2003)。この渦型歯状器官はAnaspidsよりやや遅れて出現した無顎類のThelodontsの咽頭部に既に見られる(Johanson & Smith 2003)。この仮説では、咽頭部の渦型歯条器官が口中で前方に進出し、顎部に達して歯になったとしている(Zerina & Smith 2005)(Smith 2003)(Smith & Johanson 2003)(Smith & Coates 1989)(McCollum & Sharpe 2001)(Burrow 2003)。Loanellia, Sheilia, Logjiid といったThelodonts類 の咽頭部における融合した渦型歯状器官は、詳細研究はまだであるが、無顎類と顎口類の中間形態の可能性があるとしている(Gillis & Donoghue 2007)。
軟骨魚類、棘魚類、硬骨魚類、さらには現生の鰓を持つ多くの魚類にも渦型歯状器官と同様の器官が存在している(McCollum & Sharpe 2001)。現生種の発生初期の観察から渦型歯状器官は内胚葉由来とされ、従ってこの仮説だと歯は内胚葉由来の器官となる(Fraser et al. 2009)。
なお、板皮類は歯を持たないとされているが、濾過摂食方式のCowralepisをはじめ多くの板皮類は咽頭部よりもさらに奥の鰓のあたりに歯状器官を持っている(Johanson & Smith 2003)(Carr et al. 2009)。そしてデボン紀中期に出現した板皮類のArthrodiraの系統では咬合部に新たに歯を獲得している(Zerina & Smith 2005)。このArthrodiraの歯はinside-out hypothesisに基づいて、他の魚類とは独立に咽頭部の奥、喉頭部にある歯状器官から平行進化したと解釈されている(Smith 2003)(Zerina & Smith 2005)(Carr et al. 2009)。
outside-in hypothesisとinside-out hypothesisどちらに妥当性があるか議論が継続中である。今のところ最初の仮説、outside-in hypothesisの方が第2の仮説inside-out hypothesisよりも有力視されてきたようであるが、いずれの仮説にしても歯の起源は顎の獲得以前の無顎類の時代まで遡ることになり、歯と顎は別個に進化したことになる。
さらに上述のoutside-in hypothesisやinside-out hypothesisに対して、初期の脊椎動物の歯には外胚葉由来の歯と内胚葉由来の歯といった区別はは無かったとする新たな仮説も提案されている(Donoghue & Sansom 2002)。
一方、内胚葉だけでは歯は形成されないこと、咽頭部の歯状突起が歯特有の3層構造を持たないこと、及び鰓を持つ魚類のみに咽頭部歯状器官が見られ、鰓が閉じた初期四肢類では既に咽頭部歯状器官は消失しているといったことから、modified outside-in hypothesis が提案された(Huysseune et al. 2009)(Tucker & Sharpe 2004)。この仮説では、図3に示すように、皮骨由来の歯状突起はneural crest誘導の間充織との相互作用により口から口中に侵入するとともに、咽頭部歯状器官が鰓から内胚葉上皮に進入し、さらに前方に進出して歯を形成したとしている。即ち、歯形成に際しては内胚葉由来の組織と外胚葉由来の組織との物理的な接触が関与している。

歯類は赤、外胚葉は青、内胚葉は黄でしめす。A:無顎類 BとC:顎口類 Cでは外胚葉は内部まで入り込んでいて、内胚葉を覆っていると仮定 From Huysseune et al. 2009
このmodified outside-in hypothesisは、最近報告された有尾両生類の歯発生観察からも支持されるようである(Soukup et al. 2008) (Koentges 2008)。この実験では、外胚葉と内胚葉のどちらも歯になりえることが確認されている。外胚葉のほかに上皮:epitheliaを形成しエナメル層形成に関与する内胚葉、象牙質と骨を形成する神経堤から誘導される間充織という3つの組織が歯形成には関与している。従ってoutside-inやinside-outといった従来仮説はあまり重要なことではなく、むしろ神経堤:neural crestの機能が歯の形成を決めていると結論付けている。即ち、歯形成の主役はneural crest cells であり、歯はただ一度だけ、咽頭内のどこかで、neural crest signal によって駆動されて進化したものであるとしている。

脊椎動物(上)とaxolotl(下) の口腔部における歯形成形態と歯の由来となる胚の比較。緑部は外胚葉:ETC(ectoderm)、赤部は内胚葉:END(endoderm)。大部分の脊椎動物の歯は外胚葉由来であり、外胚葉陥入内に発生する。有尾両生類の歯も外胚葉陥入内に発生するが、外胚葉と内胚葉の両方に由来する。 From Soukup et al. 2008
また、最初に歯を獲得したとされるコノドントが皮骨を持っていなかったことから、咽頭部歯状器官が歯の原型であると考え、inside-out hypothesisを元に、遺伝子解析に基づき顎の形成も加えた新たな仮説が最近提案されている(Fraser et al. 2009)。シクリッド等の発生において、顎と歯、及び咽頭部に形成される咽頭顎と咽頭歯の遺伝子発現を調べた。この仮説では魚類の歯・顎の進化は2段階を経ているとしている。500Myaよりも以前の無顎類の時期に旧歯形成遺伝子ネットワークにより、咽頭後方に外胚葉由来の歯状器官を形成した。顎口類になるときに前方咽頭弓からなる旧顎:oral jawを形成する遺伝子ネットワークに旧歯形成遺伝子ネットワークを加え、旧顎に歯を形成した。この過程で旧歯形成遺伝子ネットワークのhox遺伝子を含むいくつかの遺伝子が機能しなくなった。肉鰭類と条鰭類の分岐後、肉鰭類の系統では顎口類になったときに獲得した旧顎と旧歯を形成する遺伝子ネットワークを改変して引き継いだが、この過程で咽頭部の歯状器官を失った。一方条鰭類の方は、分岐後かなり後の真骨魚になってhox遺伝子が機能する内胚葉由来の新顎部:pharyngeal jawを咽頭部に形成し、ここに旧歯形成遺伝子ネットワークを使って歯状器官を進化させた。
肉鰭類に分岐したときに咽頭部の歯状器官を失ったことは、陸棲化に際して必要になる呼吸器系と消化器系を分離する咽頭蓋を形成する空間を咽頭部に作り出したと思われる。
初期硬骨魚とされるAndreolepisとLophosteusは歯を持っていたが、軟骨魚類と同様に歯列が後方から前方へと順次送り出されて抜け替わる方式であったとされる(Botella et al. 2007)。一方、現生硬骨魚類や四肢類は乳歯が抜けて新しい歯が同じ場所から生える方式で歯の抜け替えを行っている(Huysseune et al. 2007)。即ち、軟骨魚類と硬骨魚類の共通祖先は軟骨魚方式で歯列を入れ替えていたが、硬骨魚に分岐して後に個別の歯を入れ替える方式に変ったと考えられる。前者の方式はinside-out hypothesisの機構と関係があるのかもしれない。初期硬骨魚類から肉鰭類と条鰭類が分岐する直前に硬骨魚類の系統は神経堤が関与することで歯の交換方式が変ったと考えられる。歯の交換方式が変ったことの進化的な意味は不明である。
顎に歯を獲得し肉食化したことが顎口類の進化を加速するとともに、多くの無顎類を絶滅に追い込んだと考えられてきた。しかし、顎口類が歯を獲得してもシルル紀の間は無顎類が生態系で優勢であり、顎口類が繁栄したデボン紀になっても共存していた。
無顎類は、海底を漁る濾過摂食方式を採用していたが、草食性のHeterostracanを除くと、多くは肉食性であったとされる (Purnell 2002)。そして無顎類の恐らく総てが、所謂歯を持たないが咽頭部歯状突起を持っていたと考えられる。
さらに板皮類は歯を持たなかったが、デボン紀において非常に栄えた。そしてデボン紀後半に出現した歯を持つ板皮類Arthrodiraの1種Cowralepisは濾過摂食方式であった(Carr et al. 2009)。
従って、従来言われてきたように顎と歯を獲得して肉食化したわけではないし(Purnell 2002)、歯を獲得したからといって生態系で圧倒的に優位になったというわけでもないようである。歯を持ち肉食化した顎口類が無顎類を滅亡に追いやったということではないようである。
エウステノプテロンやイクチオステガは肉鰭類の系統に見られる迷路歯を発達させているが、陸棲化後では一部の両生類を除き新四肢類ではこの種の歯は無くなっている。エナメル質と象牙質が複雑に絡んだ迷路歯構造は、歯を強化する機能があったと考えられる。即ち、迷路歯は噛み砕くのに役立つと思われる。しかし、当時の肉鰭類や初期四肢類は餌を丸呑みしていたので、このような構造の歯を持っていた理由は不明である。また迷路歯が陸棲化に伴って急に消失した理由も不明である。
噛み砕くことが出来るようになったのは、機能の異なる歯を持つようになった哺乳類になってからである。また、上下の歯同士の噛み合わせが出来るようになったのは300Myaの羊膜類の系統であり、これにより植物食が可能になったとされる(Reisz 2006)。
まとめ
脊椎動物の歯の起源は無顎類の段階で形成された歯状器官であり、顎形成とは独立に先行して進化した。顎を獲得した顎口類でもその初期段階では顎に歯を持たなかったが、450Mya頃には歯形成の遺伝子ネットワークを顎形成の遺伝子ネットワークに組み合わせ、顎に歯を持つようになった。
顎に歯を持つようになった進化の過程に関して、皮骨性の歯状器官が顎部に流用されたとするoutside-in hypothesisや咽頭部に形成された歯状器官が顎部に移動したとする inside-out hypothesis、二つの仮説を折衷したモデルなどいくつか提案されているが、まだ結論を得ていないようである。
陸棲化に際して歯に見られる大きな変化は、一部の両生類を除いて迷路歯が消失したことであるが、機能的な面では大きな変化は無かったと思われる。