 魚が陸上を歩くまで
魚が陸上を歩くまで
第3章―6 循環系
肺はいつ、なんのために獲得されたのか
ダーウィンは、陸棲脊椎動物の肺は魚類の持つ鰾から進化したものに違いないと考えたが、この説はかなり長いこと信じられていた。20世紀後半に硬骨魚類の進化の詳細研究が進み、肺や鰾、消化管を形成する上皮細胞や筋肉組織の比較(Clack 2007)、及び分子解析からも肺と鰾の関連性が明らかにされている(Zheng et al. 2011)。しかし、鰾が肺へと進化したのではなく、魚類は最初に肺を獲得し、肺獲得後に出現した条鰭類の系統で肺が鰾に変わったことが明らかになった。また、肉鰭類では陸棲化を進めた系統において肺機能の向上が図られた(Perry et al. 2001)(Liem 1988)。
肺形成の機構としては、肺が鰓に見られる円柱状細胞:pillar cellsを持たないので鰓から進化したのでなく、咽頭部の消化器上皮に多数の血管が集まった嚢:pouchを発達させ、これが鰓の機能を補完して肺の機能を持つようになったとされる。
しかし、多くの条鰭類では鰾が消化管の背側に、一方、肉鰭類や四肢類では肺が消化管の腹側にあり、鰾と肺が相同器官であるとする説では説明できないとされている。そのため、肺から鰾が進化した、あるいは鰾と肺が相同器官であるとする結論には若干の疑問が持たれているようである(Daniel et al. 2004)。
魚類が肺を持つようになった原因に関する様々の仮説が提案されてきている(Liem 1988)。よく知られている仮説として「涸れ池仮説」や「低酸素仮説」があるが、このほかにも「心筋への酸素供給仮説」(Farmer 1997)(Farmer 1999)、さらには現生のコイやニシンなどでは鰾が聴覚の向上に寄与していることから「聴覚機能仮説」など種々の提案がなされてきた。
これらの説の中で長いこと支持され最もよく膾炙しているのが、所謂「涸れ池仮説」である。二十世紀初頭デボン紀の化石の多くは赤色砂岩層:Old Red Sandstoneから出土していた。地質学者バレル:Barrell は1916年にデボン紀後期の地層に見られる赤色砂岩層は泥の中に含まれる鉄分が空気に曝されて酸化したために形成されたものであり、乾燥した環境で形成されると考えた。従ってデボン紀に四肢類の祖先種が棲息していたのは熱帯の雨季と乾季の差が激しい地域であった結論した。そして乾季に干上がった小川や湖といった淡水域で生き延びるために肺を獲得したとする「涸れ池仮説」を唱えたとされる(ジンマー)。デボン紀中期、385Myaの地層から出土した淡水性のハイギョ化石が空気呼吸していたことが示され、「涸れ池仮説」を支持するものとされた(Clement & Long 2010)。実際解説書や専門書でもこの仮説を採用しているものが多い。
しかし、その後化石記録から当時の熱帯地域で脊椎動物の陸棲化が進んだことは確かであるが、赤色砂岩層は必ずしも乾燥気候や淡水域で堆積するとは限らないことが明らかになり、「涸れ池仮説」に疑問が持たれた(クラック)。
それに代わりに熱帯における淡水は暑さのために水の溶解酸素量が低下するという説や暖かくて富栄養の淡水は酸素欠乏になり易いという説が出され、乾季や干上がりに関係なく熱帯の酸素欠乏を起こしやすい淡水域の環境が肺を発達させたとする「酸素欠乏仮説」が出された(ローマー)。「涸れ池仮設」に代わる「酸素欠乏仮説」でも淡水域で肺が獲得されたとしている。
一方、熱帯地方の淡水湖は乾季でも酸欠状態にならないことが明らかにされている(Grigg 1965)。さらに、化石の詳細解析により、肉鰭類は海岸の潟に棲息していたことが明らかになった(ジンマー) (Thomson 1969)。1990年代になると同位体分析技術の向上により、シルル紀やデボン紀の魚類の多くが棲息していたのは沖合いの海または汽水域であることが明らかになった。そして当時の海の少なくとも表層部は酸素不足の状態になかったとされている(Bray 1985)(ザラーシヴィッチ)。
これらの事実から、一般に受け入れられている淡水において肺が獲得されたとする説に代わって、汽水域で肺が獲得されたとする説が提案された。富栄養環境の潟は酸素不足に陥りやすいので肺を獲得したとする説や浅瀬や潟といった環境で次第に曝気状態に適応していく過程の中で肺を獲得したとする説が提案された。
最近化石の詳細分析結果を大気中酸素分圧の変動に関連付け、大気中酸素分圧が魚類の肺機能獲得と関係があるとする報告がなされた(Clement & Long 2010)。375Myaの浅海の礁に棲息していたハイギョ化石の頭胸部骨格形状からair-gulpingで空気呼吸をしていたと判断した。そして デボン紀中期に大気中酸素分圧の低下があったが(Ridgwell & Zeebe 2005)、一方魚類はより高い新陳代謝を必要としたため空気呼吸の能力を獲得したと考えた。このような呼吸法と肺の獲得はハイギョでは淡水域でも海水域でも起こったし、四肢類様魚類Gogonasusu等でも同様のことが起こったと結論した。即ち、380~370Myaの大気中酸素分圧が低い環境で淡水性、海水性を問わずハイギョや四肢類様魚類が肺を獲得したと主張している。これは「酸素欠乏仮説」の範疇になり、少なくとも肉鰭類が肺を獲得したことは説明可能である。しかし、低酸素時代だったとされるデボン紀中期よりもあるか以前の420Myaには条鰭類が肉鰭類と分岐していたとされており、条鰭類の肺の獲得が説明できない。
また、熱帯の干満域に棲む現生のハゼ類のような海生両性魚類は初期四肢類に似ており、海生両性魚類をデボン紀の魚類上陸、あるいは肺の獲得と関係づける新たな説が出ている (Ip, Y. K. et al. 2004)。しかし、干満域での選択圧では完全な陸生化のための呼吸系を得ることは出来ないとされる(Graham & Lee 2004)。いずれにせよ現生のハゼ類は肺・鰾を獲得後に半陸棲化した魚類であり、必ずしも古生代の肺・鰾を持たない状態から新たに肺を獲得した魚類との比較・類推は妥当ではないだろう。
第1章や第2章で既に述べたように、最近の化石情報(Blieck & Turner 2003)の増加と分子生物学の発展(Donoghue et al. 2003)にともない、硬骨魚類から420Myaには既に肉鰭類と条鰭類は分岐しており、さらに条鰭類では肺が鰾に進化したことが、即ち肺と鰾とは相同器官であることが分子解析から明らかにされている(Zheng et al. 2011)。従って、肺獲得に関するこれらの仮説の多くは初期硬骨魚類が420Myaより以前に肺を持っていたことを説明できない。
脊椎動物における肺の獲得は肉鰭類と条鰭類の分岐より前であり、シルル紀中期から後期にかけて棲息していた初期硬骨魚類の時代と考えられた(Zhu et al. 2006) (Perry et al. 2001)(Liem 1988)。
さらに最近の棘魚類化石の詳細な調査から、棘魚類も初期的な肺を持っていることが示された(Nelson 2006)(松井 2006)。棘魚類は、肺以外にも、三半規管と3個の耳石、骨性の鰓蓋、前鼻孔と後鼻孔が分離、神経頭蓋腹面に噴水溝(spiracular groove)、神経頭蓋の耳殻部と後頭部を分離する耳―後頭裂溝(otico-occipital fissure)、神経頭蓋の基底の前後の化骨部を分離する基底裂溝(ventral fissure)、頭部に大きな皮骨要素等があり、硬骨魚類と多くの共通派生形質を持っている。
こういったことから硬骨魚類と棘魚類は姉妹群とされ、硬骨魚類と棘魚類の共通祖先と位置づけられる真口類: Perfect mouth:Teleostomiの存在が予想される(松井 2006)。従って脊椎動物における肺の獲得は、硬骨魚類と棘魚類の共通祖先とされる真口類にまで遡ることになる。真口類は軟骨魚類にやや遅れるがほぼ同じ頃、450~430Mya頃に出現していると考えられる。従って、脊椎動物の肺は430Myaまでには既に獲得されていたことになる。
魚類が420Myaより以前に肺獲得したことは確かと考えられるが、何ゆえ肺が獲得されたのかはもちろん淡水域、海水域のいずれで肺獲得が為されたのかに関しても明確な結論は出ていない。
化石記録ではオルドビス紀にはすでに顎口類が出現したが、これは分子時計による予想とほぼ一致している(Blieck & Turner 2003)。顎口類出現後もしばらくは無顎類が支配的であり、出現後20Myほどした430Mya頃になって顎口類は放散を始めた。肉鰭類と条鰭類は420Myaまでには分岐したとされており、従って顎口類が放散を始めた時期は軟骨魚類や棘魚類、板皮類といった顎口類ばかりでなく、初期硬骨魚類もすでに出現していた可能性がある。
初期硬骨魚類と考えられる422~420Myaに生息していたAndreolepisやLophostensは化石が不完全であるが、その体形や鱗などから板皮類のような底生ではなかったと考えられる。さらに419Myaに棲息していたGuiyuは初期硬骨魚類から分岐直後の肉鰭魚類と考えられ、その流線型の洗練された体型や口吻が前方にあること、鱗の形状などから遊泳性であったと考えられる。これはGuiyuの祖先である初期硬骨魚類が遊泳性であること示唆している。
背骨の充分な骨化はペルム紀になってからであるが、初期硬骨魚類では、完全ではないにせよ背骨の硬骨化が見られ、これにより筋が付着する体幹部が強化された。さらに鱗を軽量化するとともにペグとソケットを設け、鱗同士の結合力を高めて体部の剛性を強化している。初期硬骨魚類における硬骨化と鱗へのペグとソケットの付加は体幹筋肉の強化、即ち遊泳力向上への適応である。鱗の小型化・軽量化も遊泳力向上への適応であったと考えられる。また、先の3章-3 「鰭と四肢」で見たように初期硬骨魚類は機動性と遊泳性の向上のために、胸鰭筋肉原基となる先駆筋芽細胞を側板中胚葉から拡散によって必要部位に運ぶ方式を進化させることで対鰭の可動性を増し、肩帯骨格の強化により胸鰭筋肉の増強を可能にしている。さらに口吻の位置や外形からも、初期硬骨魚類は遊泳型の魚類であったことが示唆される。
第2章で見たように棘魚類は遊泳性の魚類であり、初期硬骨魚類も遊泳型の魚類であったと考えられる。従って、棘魚類と初期硬骨魚類の共通祖先である真口類の化石報告の有無は不明であるが、真口類も遊泳性の魚類であったと予想される。
真口類が出現したと予想されるシルル紀初期から中期にかけて、海域では遊泳力を持った肉食性の軟骨魚類が徘徊していたので、真口類が生き残るためには高度な遊泳力を備える必要があったであろう。
当時鰓だけで呼吸を行っていた無顎類や軟骨魚類は、現生魚類と同様鰓から酸素を血液中に取り込み体組織に供給し、酸素の欠乏した血液を心臓に戻していたと考えられる。従って、長時間の過激な運動により心臓の筋肉が酸素欠乏状態になりやすい構造になっている。一方、現生の古代魚アミアのように肺を持つ魚は、鰓→体→心臓→鰓という普通の魚の血液循環ルートのほかに肺→心臓→鰓の血液循環ルートを持つ。このような背景をもとにファーマーは心臓に酸素を供給するために肺を獲得し、これによって長時間の高速遊泳が可能になったという説を提案した(Farmer 1997)(Farmer 1999)。図1に条鰭類、アミア、及びハイギョの心肺系を示す。アミアやハイギョでは肺を経由した酸素の多い血液が心臓に直接送り込まれるようになっている。
真口類が肺を獲得したと思われる430Mya頃は、酸素分圧が上昇していたとされる時代であり(Berner 2006)、従って海水中酸素濃度も高い値を示していたと考えられる。即ち、活動的な顎口類はより多くのエネルギーを必要とするため、酸素分圧の上昇が顎口類放散のトリガーになったと推論される(Qu et al. 2010)。しかし、シルル紀には深海が何回か無酸素状態になったとされる(Zalasiewcz 2010)。従って、遊泳性の魚類の棲息域は沿岸部の浅海域か、外洋の海表面域に限られており、遊泳性に優れた捕食性の魚類同士の厳しい競合があった時代であったと思われる。

b. 条鰭類:酸素の少ない血液を心臓に供給 c. アミア:初歩的空気呼吸型魚類:酸素の多い血液と酸素の少ない血液を混合して心臓に供給 d. ハイギョ:空気呼吸型魚類:酸素の多い血液を心臓に供給 From Farmer 1999
こういった環境に出現した真口類にとって、遊泳力の持続性を保証するのが、心臓への酸素供給を可能にする肺と血管系を含めたシステムの獲得であったと考えられる。こういったことからファーマーによる「心臓への酸素供給のために肺を持った」という仮説が最も妥当性があるように思われる。即ち、魚類の肺は、表層部を遊泳する真口類の時期の430Mya頃に獲得されたと考えられる。
現生軟骨魚類の個体発生の研究から、軟骨魚類がかって肺を獲得したことはないと考えられている。板皮類も肺を持っていなかったとされる。しかし、デボン紀後期に繁栄した板皮類のBothriolepis は肺様組織を持つとの化石報告があるが(Perry et al. 2001)、他に同様の報告が無いので確実でない。もし底生の板皮類が肺を持っていたとすれば、遊泳性の真口類とは独立に肺を進化させたと考えられる。デボン紀後期は低酸素環境化であったとされ、この場合の板皮類における肺の獲得は、低酸素への適応だった可能性があり、従来の肺獲得に対する「酸素欠乏仮説」の範疇になろう。
現生ハイギョは遊泳性ではないが肺を持つ。また鰓を経由せず、心臓から直接全身を循環する回路の形成が見られる。このようなシステムは鰓が機能しない乾燥下における長期生存のため、他の四肢類とは独立して進化したものと考えられる。即ち、ハイギョの肺は真口類の時期に獲得されたが、肺・心臓・体部の循環系は「涸れ池仮説」に基づいて形成された可能性がある。
鼻孔の獲得
陸棲脊椎動物は鼻孔を呼吸と臭覚の器官としているが、魚類の鼻孔は臭覚器官であり、本来鼻孔は呼吸と関連が無かった。軟骨魚類は腹側に硬骨魚類は背側に鼻孔を持つ傾向が見られるが、いずれの鼻孔も2対で4つの孔から構成される。即ち前方に前鼻孔と呼ばれる左右一組の流入孔と後方に後鼻孔と呼ばれる左右一組の流出孔から成っているのが基本形である(Fishelson & Baranes 1997)。
左右に対となった鼻孔を持つ脊椎動物は、無顎類ですでに出現している。無顎類では2対の鼻孔を持つ種類と正中線上に1対の鼻孔を持つ種類がいる。前者には翼甲類 or 異甲類:プテラスピス等が、後者には頭甲類:ケファラスピス等、欠甲類:ヤモイチウス等、さらに現生ヤツメウナギ類(鼻孔は盲嚢状の鼻嚢に接続)、現生ヌタウナギ類(鼻孔は咽頭部に開口)がある。現生ヤツメウナギやヌタウナギが正中線上に鼻孔を持ち、魚類や陸棲脊椎動物が左右対となった鼻孔を持つことから、前者の方が原始的に見える。しかし、正中線上に1対の鼻孔を持つ頭甲類、欠甲類より先に出現した翼甲類は2対の鼻孔を持っており、2対の鼻孔の方が進化した形態とは必ずしもいえないようである。いずれにせよ、2対の鼻孔は無顎類の翼甲類の時期に獲得した形態といえる。
現生魚類では2つの前鼻孔と2つの後鼻孔を持ち、現生四肢類では2つの前鼻孔を持つが後鼻孔は無く、鼻腔になっている。395Myaに棲息していたKenichthysは内鼻孔を持っているが(Zhu & Ahlberg 2004)(Janvier 2004)、まだ歯列のあたりに留まっている。またOsteolepoform :Osteolepis and Eusthnopteron 等は2つの鼻孔と口蓋に鼻腔様の空間を持つ。
内鼻孔の獲得は底性化への適応と考えられる。即ち、底性化したことにより従来の前鼻孔と後鼻孔という構造では鼻孔内の水流を確保出来なくなった。そのため内鼻孔を設け、鰓蓋を動かすことによって鼻孔内の水流を確保し、嗅覚器官として機能するようにしたと思われる。
シーラカンスは内鼻孔を持っていないが、シーラカンスが肉鰭類の系統から出現したのは肉鰭類が分岐してまもなくであるとされる。またシーラカンスに続いて肉鰭類の系統から出現したハイギョでもその祖先と考えられる400MyaのDiabolepisは内鼻孔を持たず4つの鼻孔を持っている。内鼻孔の形成が底生化と関係すると考えられるので、シーラカンスやハイギョが内鼻孔を持たないことは、初期硬骨魚類から分岐した直後の肉鰭類が遊泳性であったことを示唆している。
一方、現生ハイギョは前鼻孔を持ち、後鼻孔の代わりに内鼻孔を持っているが(Young 2010)、現生ハイギョの内鼻孔は、鼻関連神経系、骨格、側線等の観察から、四肢類様魚類とは独立に進化したものであることが明らかにされている(Long et al. 2008)。
後鼻孔、内鼻孔、鼻腔の相関に関していくつかの議論が交わされてきたが、そのなかで有力な2説は、(ⅰ)後鼻孔は鼻腔の祖先型であるが、ハイギョの内鼻孔とは無関係 (ⅱ)鼻腔は新奇に獲得されたものであり、後鼻孔は消失したか、または涙管(tear duct)になった、というものである。化石の詳細分析とエヴォ・デヴォにより内鼻孔が鼻腔に対応するものであり、さらにハイギョの内鼻孔は鼻腔と相同器官であることが確認された(Zhu et al. 2009)。従って、後鼻孔→内鼻孔→鼻腔という変化は、肉鰭類の進化に共通するものと考えられる。しかし、鼻孔を臭覚器官としてだけでなく、呼吸のための器官として使うようになったのはかなり後のことで、完全な陸棲化を果たした四肢動物になってからであろう(クラック)。
2心房化
現生哺乳類では、心房や心室、心臓周辺の血管系の形成にはNkx2-5/Tbx5/Mf2C/Hand2 などがプログラムの活性化を、Nx2-5/Tbx2/Tbx3がプログラムの抑制を、さらにAnf, Cx40, Cx43が形成を担っている(Antoon et al. 2003)(Koshiba-Takeuchi et al. 1988)。また、魚類から四肢類に到るまで心房側(inflow side)ではtbx-5, hrt1, raldh2, couptf-Ⅱ, amhc1といった遺伝子が、心室側(outflow side)ではIrx-4, mlc2-v, hrt-2といった遺伝子が関与している(Simoes-Costa et al. 2004)。魚類の心臓から両生類、羊膜類の心臓へと進化する過程において、これら転写因子や制御遺伝子等が共通して関与し、その発現の様相が変化したと考えられる。
心臓の形成において、魚類の蠕動運動で機能するパイプ形状をした心臓が両生類の同期運動をするループ状の空間を持った心臓へと進化した。その進化の過程は不明だが、左心房の形成が魚類から両生類への進化の過程で起こっている。心臓形成時に左側で発現するPitx-2遺伝子がこの進化に関与していると思われる。この結果心房隔壁形成と静脈洞を囲む心筋形成に加え、心臓への入路が肺からと体組織からの2系統になった(Antoon et al. 2003)。
底生型の肉鰭類の段階では、摂食行動は待伏せ方式であり、活動的な行動は一時的な摂食行動時に限られているため体細胞への多量の酸素供給は必須ではなかったであろう。従って、395Mya頃に底生化したKenicchthysの心臓は、硬骨魚類の段階にあったと思われる。底生化した肉鰭類はその後浅瀬に進出したと考えられ、泥濘中や草木の茂る中を移動するようになった。水中を遊泳する移動方式や水底で待伏せしているよりも泥濘中を這い回る移動方式の方がよりエネルギーを必要とする。肉鰭類は潮汐域に進出したことにより、体組織への多量の酸素供給がより必要になり、その適応として2心房化が起こったと考えられる。従って、2心房化に向けた進化は肉鰭類が潟などの浅い潮汐域に移行を開始した時期、即ち385Mya頃に始まったと思われる。
「ローマーの空隙」が終わった直後、340Mya頃にはかなり活動的と思われる体形の両生類が出現しており、2心房化を完成し、弁機能、および心臓と近傍の血管の拍動差により、1心室における肺からの酸素の多い血液と体からの酸素の少ない血液の混合を抑えることで、効率の高い心臓を進化させていたと考えられる。即ち、2心房化とそれに伴う循環系は385Myaから340Myaにかけて時間をかけて形成されたものであろう。
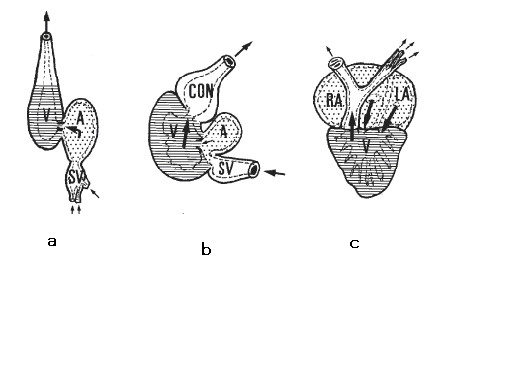
C:サメ、D:硬骨魚類、E:両生類 V:心室 ventrecle A:心房 atrium SV:静脈洞 sinus venosus CON:動脈円錐 conus From 大石正道2008
現生シーラカンスは1心房であるが、これは肉鰭類の系統が潮汐域に進出するよりはるか以前のまだ遊泳性であった時期に肉鰭類から出現したことを反映していると思われる。また 現生ハイギョに2心房化の初期状態が見られることは、長期の乾燥に、従って鰓が長期にわたって機能しないことへの対応と考えられ、四肢類の系統とは独立に進化したものである。
条鰭類と肉鰭類のいずれも腎臓からのナトリウム排出を制御する副腎皮質ホルモンであるアルドステロンの1種CYP11Bを保持している。そして肉鰭類は条鰭類との分岐後で、しかも370Myaより前にCYP11B2を遺伝子重複で獲得したことが遺伝子解析から結論された(Colombo et al. 2006)。現生ハイギョもCYP11B2を持っており、このホルモンは低酸素環境への対応のためとされる。従って、肉鰭類は370Myaまでのどこかの時点で低酸素雰囲気下に生息していたと考えられる。
最近のモデル計算によると380Mya頃に大気中酸素分圧が非常に低下したとの結果を得ている(Benton 2010)。低酸素化は、肉鰭類が浅瀬や潟へと進出し、肺呼吸への依存性を高め、陸棲化へ向かう契機になったと考えられる。この時期はアルドステロンから予想された370Nya以前の低酸素環境の経験と一致している。
呼吸系の系統による違い
四肢類は消化管の腹側に対になった肺を持っており、一方条鰭類の多くは消化管の背側に鰾を持っている。しかし、現生ポリプテルスは消化管の腹側に1対の肺を持っているが、軟質魚類に分類される現生チョウザメは、ポリプテルスとほぼ同じ時期に条鰭類から分岐したとされているが、背側に鰾を持っている。
鰾と肺が相同器官であるとする説が有力であるが、こういった肺・鰾の違いやその位置が異なる理由を説明出来ないため、相同器官とする説に疑問が持たれている(Daniel et al. 2004)。初期硬骨魚類を祖先とする現生の脊椎動物を含めた概略の分岐系統樹に肺または鰾の保持形態、保持位置、呼吸方式をまとめて図3に示す(Perry et al. 2001)。
発生初期に出現する7番目と8番目の咽頭嚢から肺が、6番目の鰓弓から肺動脈が形成されたのは、初期硬骨魚類の時期と考えられる。肺が消化器系から独立し鰾になったのは真骨魚類の時代になってからである。従ってチョウザメの肺から鰾への進化は、真骨魚類の肺から鰾への進化とは独立したものと考えられる。板皮類の系統でも硬骨魚類の系統とは独立に肺を持った可能性がある。しかし、肺様器官らしきものがあるとされる化石報告はただ一例であり、板皮類が肺を持っていたことに関しては確実ではない。

肺位置の系統による違い
肺の位置と呼吸法を入れた図3の脊椎動物の進化系統図を、肺/鰾位置の違いに注目して書き変えたのが図4である。軟質魚類のチョウザメや全骨魚類のガーやアミア、さらには真骨魚類を含めてすべて肺/鰾は消化器系の背側にある。一方、初期段階の条鰭類から分岐したとされるポリプテルスでは肺が消化器系の腹側にあり、陸棲脊椎動物でも肺は消化器系の腹側にある。またほぼ同じ時期に相前後して肉鰭類の系統から分かれたシーラカンスの鰾は消化管の背側にあり、ハイギョの肺は消化管の腹側から出て消化管をはさんで両側に対となっている(Long)。
条鰭類と肉鰭類の共通祖先である真口類乃至は初期硬骨魚類が肺を獲得していたことは確かであるが、こういった肺ないしは鰾の位置が異なることを説明できない。

現生種における肺や鰾の位置を示す。V:消化管の腹側に肺、または鰾を持つ系統 D:消化管の背側に肺、または鰾を持つ系統 N:肺も鰾も無い系統
空気呼吸をする必要が生じて最初に消化管の一部が嚢状になり、そこに細い血管が密に張りめぐらされて肺の原型が出来たと考えられている。口蓋と消化管をつなぐ咽頭部の口蓋に近い前方の部分は、その発生段階において咽頭弓が左右に5対ほど並んでいる。これら咽頭弓は頭部や頸部の種々の器官形成に関与しており、従って、咽頭部のこの部分に肺の原型となる嚢を形成することは不可能であったと考えられる。そのため咽頭部の一番奥の喉頭部に肺の原型となる嚢が形成された。嚢状の器官が消化管に形成されるのは腹側、背側いずれの側も優先性は無く、どちらに形成されるかは確率的に等価と考えられる。即ち、初期硬骨魚類において消化管の腹側に肺の原型を持つ魚類も現れたであろうし、背側に肺の原型を持つ魚類も現れたであろうと予想される。
遊泳性に優位性を置く魚類では、形成された肺原型が腹側にあるよりは背側にあるほうが、浮心が上になる傾向が強まるのでより安定した遊泳が可能であり、選択圧がかかったと考えられる。即ち、条鰭類は390Mya頃から放散を始めたが、現生条鰭類のほとんどは背側に肺/鰾を持つことから、恐らく放散する前に条鰭類の系統では肺の位置が背側に固定したと考えられる。
一方、遊泳をほとんどしない底生の魚類では、背側、腹側いずれでも優位性は無かったであろう。ポロプテルスは、条鰭類が放散を開始する前に条鰭類の系統から分かれた(Inoue et al. 2005)。ポリプテルスの祖先は底性であったために腹側に肺の原型を固定しても特に問題は無かったであろう。ポリプテルスよりやや遅れた時期に出現したチョウザメなどの軟質類が分岐したのは、条鰭類の系統で肺位置が固定された後であったと考えられる。
底生に移行した肉鰭類は395Mya頃に内鼻孔を口蓋先端部に持ったが、陸棲化に伴い内鼻孔は消化管の背側に沿って次第に奥の方に移動したとされる。
しかし、イクチオステガの段階でも内鼻孔は口縁近くにあり、呼吸には使われず、臭覚器官として機能していた。イクチオステガは鰓を持っており、扁平な口吻であり、従って鰓呼吸と口から空気を飲み込む肺呼吸を併用していた。イクチオステガとほぼ同じ頃浅い潮汐域に棲息していたチュレルペトン:Tulerpetonは鰓を持たなかったが、扁平な口吻を持っており、口から空気を飲み込む肺呼吸を行っていたと思われる。従って、チュレルペトンの段階でも内鼻孔は口吻近くにあったであろう。
「ローマーの空隙」の時期に出現したPederpesやWhatcheeriaは陸上歩行が可能な四肢を持つ、体長1メートルほどの両生類であった。これら新四肢類は、幅の狭い頭骨を持っていることと筋肉の付き方から、口から空気を飲み込むのではなく、肋骨を使った肺呼吸を行っていたと考えられる。即ち、PederpesやWhatcheeriaの鼻は呼吸器官としても機能していたと考えられる。従って、内鼻孔はイクチオステガのように口縁近くではなく、喉頭部まで下がってきていたと思われる。これら新四肢類では喉頭部の背側に内鼻孔が来たため喉頭部の腹側に肺原型は形成せざるを得なかったと考えられる。
即ち、四肢類において肺の位置が消化管の腹側に固定されたのは、初期四肢類ではなく、新四肢類になったこの時期であったと思われる。
真口類や初期硬骨魚類では、肺は消化管の腹側、背側どちらに形成されるかは、確率的に同じであり、腹側に肺を持つ魚類も背側に肺を持つ魚類も出現していたと思われる。その後遊泳性に優れた多くの条鰭類は背側に肺を固定する方向に選択圧が働いたと考えられる。一方、四肢類は内鼻孔を持っていたために腹側に肺を固定せざるを得なかったと考えられる。
現在知られている最古のハイギョは417~416Mya頃棲息していたStyloichthysやPorolepisである。一方、知られている最古のシーラカンスは最古のハイギョよりは新しく410Mya頃棲息していたEuporosteusであるが、シーラカンスの方が出現はやや早いとされる。いずれにしても肉鰭類と条鰭類は420Myaには分岐していたとされるので、シーラカンスやハイギョは肉鰭類が分岐してまもなく出現したことになる。
上述のような機構で肺の位置が固定されたとする仮説に基づくと、現生シーラカンスとハイギョの肺形成過程は次のようになる。現生シーラカンスの肺は背側にあり脂肪が充填された鰾として機能しており、従ってシーラカンスは遊泳性の範疇に入る。シーラカンスが分岐したころは、肉鰭類は遊泳性であったと考えられ、シーラカンスはその形態を保持してきたと考えられる。一方、現生ハイギョの肺は消化管の腹側につながっている(Long)。ハイギョは肉鰭類の系統から分岐後底生になり、背側、腹側いずれの種類も出現したであろう。底生では肺位置の腹側、背側の優位性が無いため、たまたま肺が腹側に固定された方が生き残ったと思われる。
棘魚類は最初は海域に棲息していたが、条鰭類が放散を始めると共に淡水域へと進出し、淡水域で繁栄している。恐らく遊泳性で競合する条鰭類と棲み分けたと思われが、石炭紀後期には条鰭類も淡水域に進出している。遊泳能力で劣ると考えられる棘魚類が、条鰭類の出現後も三畳紀前期まで生存し、淡水域でもお互いに5000万年もの間共存していた。このことは、少なくとも棘魚類が繁栄した河川や淡水湖は一般に海に比べて狭く浅いため、遊泳能力の違いは生存上あまり問題ではなかった可能性がある。
なお、条鰭類では肺から鰾へ進化しているが、肺が消化管から切り離され完全な鰾になったのは、ジュラ紀頃に出現した真骨魚類になってからと言われている。シルル紀前期頃に肺を獲得し、ジュラ紀になって完全に消化管から独立した鰾へと2億数千万年かけて進化させたことになる。これだけの時間を要したのは、それまでの生態系では完全な鰾が生存上必須の器官ではなかったことを予想させる。完全に消化管から独立した鰾でなくても浮力を生み出す機能は充分あったためと思われる。実際、現生の真骨魚類でもイワシ目、ニシン目、サケ目、コイ目など多くの魚において鰾が気道を通じて消化管につながっており、これらの魚類では水面から口で直接に空気を鰾に取り入れている。
条鰭類において肺から鰾へと進化した原因に関しても多くの仮説がある。肺呼吸のために棲息域は浅い水域に限られていたであろう。水面生態系から離れ完全な鰾を持つようになったのは、翼支竜や鳥類といった滑降型捕食者の出現により空気呼吸中の捕食から逃れることが原因だったとする説(Farmer 1999)、淡水域から海へと進出した条鰭類が海水中に十分な酸素が存在するので鰓だけで必要量の酸素を獲得できるようになったので肺が無用になって鰾に進化したという説(ローマー)、深海への進出に伴い空気呼吸に要するエネルギーが増加したことため肺が不用になった、などが出されている。
外洋性の魚類では浮力に振り分けるエネルギーを減らす必要があり、そのために肺を鰾に変えたとする説(Liem 1988)は、肺を持たない外洋性の軟骨魚類が肝臓に油を保持して浮力を得ていることから妥当性があろう。鰾を持っていないことは浮力のためのエネルギーを要することであり、不利であったようである。
また、外洋を高速で移動する魚類では、空気だけを口から吸込むことが難しく、肺への海水の流入が避けらなかったため肺呼吸をやめざるを得なかったとも考えられる。
脊椎動物の生理機能においては、炭酸ガスの処理能力は体液酸性度の安定にとって重要であり、酸素の欠乏よりも炭酸ガスの体内蓄積の方が制限因子になる場合が多い(クラック)。現生ハイギョでは酸素呼吸の90%を肺から採っているが、炭酸ガス排出では肺が30%、鰓が70%を占めるとされる。現生の両生類の肺は、酸素を吸収する機能を持つが、炭酸ガスを排出する機能はほとんど無く、鰓と皮膚によって炭酸ガスを排出している。一方、羊膜類の肺は酸素の吸収機能と炭酸ガスの排出機能の両方を持っている。しかし、こういった炭酸ガスの排気機能を肺がいつ獲得したのかに関しては明らかではないとされる。
鰓は構造上乾燥に対して脆弱であり、鰓の曝気が頻繁になると同時にその機能を失ったと考えられる。初期の四肢類は腹側の皮骨性鱗は残しているが、背中の皮骨性鱗を捨てている。背側の鱗をなくした結果、水面から出る部分は乾燥の危険性に曝されたが、一方、皮膚呼吸を行えるようになり、炭酸ガスを放出する機能を持てた可能性がある。とはいえ、アカントステガやイクチオステガのような初期四肢類はかなり大型であり、皮膚呼吸への依存性はたとえあったにしてもそれほど高くは無かったと予想される。
初期四肢類の出現後「ローマーの空隙」があり、この間に小型で皮膚呼吸を可能にした種類や鰓を残した種類が現生両生類の祖先となったのであろう。なお、現生両生類の祖先に関しては議論が進行中である(Marjanovic & Laurin 2007)。四肢類が鰓を消失した時点で、従って、チュレルペトンTulerpetonの段階で、肺は炭酸ガス処理機能を持ったと考えられ、鼻を呼吸器官にするよりもやや早い時期となる。鼻を呼吸器官にも適応した四肢類はPederpesやWhatcheeriaになってからと考えられる。
まとめ
棘魚類と初期硬骨魚類の共通祖先である真口類は、軟骨魚類よりやや遅れて出現したが、その体の特徴から遊泳性の軟骨魚類と競合する存在であった。そのため真口類は、肺を形成して心臓への酸素供給能力を高めることで持続的な遊泳能力を獲得した。
初期硬骨魚類から420Myaまでには分岐した肉鰭類もその初期には遊泳性を引き継いでいた。しかし、395Mya頃出現したKenichtysは内鼻孔を持ち、385Mya頃棲息していたEusthenopteronは腰帯にも四肢類的特徴を持つようになった。即ち、395Mya頃から多くの肉鰭類は底生へと変り、385Mya頃には潟などの潮汐域に進出した。頭部の扁平化は浅瀬や潟への適応形態とされており、従ってGogonasusが出現した380Mya頃に浅瀬や潟へ本格的に進出し始めたことになる。
浅瀬への進出に伴い、移動のために鰭から四肢への変遷が進行し、また、曝気が増えるのと同期して鰓機能が衰退した。さらに肺機能の向上、心臓の2心房化が始まり、循環系に適応進化が起こった。循環系に起こった適応進化は、水底、あるいは潮汐域といった水棲時期に起こったものであり、鰭から肢への適応進化と同様、その後の陸棲化の前適応である。
肺と鰾は相同であり、消化管の背側、腹側のどちらに始原肺の嚢が形成されるかは、確率的に同じである。しかし、浮心、重心といった物理的条件により、遊泳性の魚類では背側に肺を固定する方向で選択が働いた。底生性の魚類や潮汐域へと進出した四肢様魚類や初期四肢類では肺位置に関する背側、腹側の優位性はなかったと考えられる。鼻と肋骨が肺呼吸に使われるようになった新四肢類の頃に、内鼻孔が喉頭部まで下がり、腹側に肺を固定せざるを得なかったと考えられる。